
第1回研究会での発表内容
1. 趣旨説明(河合香吏)
本共同研究課題は、2005年度から「人類社会の進化史的基盤研究」として、(1)集団(2005-2008年度)、(2)制度(2009-2011年度)、(3)他者(2012-2014年度)、と主たるテーマをかえて3期10年にわたって展開してきた共同研究の第4期にあたる。また、一連の共同研究は今期でいったん閉じる方針であるため、その総括としての意義をも兼ね備えている。
一連の共同研究は当初から長期プロジェクトとして企画され、群居性の霊長類の一員としての人類が進化の過程で獲得してきた高度な「社会性(sociality)」を種の誕生における最重要の特質として着目しようとするものであった。「社会性」なるものは、精神や社会の進化と同様に、化石として形に残りにくいものであるため、ヒトの社会とヒト以外の現生霊長類の社会との比較・検討が不可欠であった。そのため、霊長類学者と人類学者との共同研究が強く求められ、両者の共同討議として進められてきた。
今期は上記一連の共同研究の総括をするにあたり、「生存・環境・極限」といったやや広いテーマを中心課題として設定した。これは、研究主題としては「生存」を主軸とし、「生存(「生きる」ということ)の環境」を、その「極限」条件ないし「極限」状態まで見据えるものとした。ここで言う環境とは、生態学的かつ社会的であるような環境を指すが、そうした環境に「生きる」ことのぎりぎりの条件ないし状態を探ることを念頭に、これまでと同じ3つの学問分野である、霊長類社会・生態学、生態人類学、社会文化人類学からこの課題に迫りたい。
これまでの一連の共同研究は、ヒトを含む霊長類が適応したり発達させたりしてきた自然的/社会的環境を「舞台」ないし前提として展開されてきたものであり、複数の同種個体がともに<生>をはぐくむ術、生態学の用語を使えば「生存戦略」だが、そうしたものを明らかにすること、そして、人類が高度に発達させてきた「社会性」(sociality)の起原と進化の機構について考察することを目指してきた。
生物としての<生>(「生存」と言っても「生命」、「いのち」と言ってもよいが)、すなわち「生きる」ということは、まずは生物学的/生態学的に論じられうるものだが、それに一元的に還元することはむずかしい。とりわけわれわれ人類を含む霊長類が群居性動物として生きる存在である以上、そこには社会的なるものが必然的な要素としてあり、他者と「ともに生きて」いかねばならないということ、そしてそれ故に、その方法(生存戦略)は、非決定論的で、フレキシブル(可変的)で、コミュニケーショナルなものになると思われる。こうした性格については、すでに「集団」、「制度」、「他者」というこれまでの共同研究においても議論され、明らかにされてきたことでもあるが、この点に議論を焦点化して、人類の社会性の進化の解明に向けて新たな視座を追究していきたい。
本研究課題ではこれまでの一連の共同研究において明らかになっていたことを基盤として、その総括として、ヒトとヒト以外の霊長類の生物学的、生態学的環境の極限、および社会性の極限を詳らかにしていくことを通じて、人類社会における<生>=「生きること」の進化史的基盤を考えていくことを目指す。
以上のように、環境の極限を明らかにするとどのような学問的貢献があるかということの、より積極的な意義についても触れておきたい。まず、進化を遂げてきた社会の各段階のすべてに常に存在する「基盤」なるものが果たして何であるのかを明らかすることから、それは、「社会論」一般に寄与することになるだろうということが挙げられる。同時にそれは、社会のさまざまな危機、たとえば、急激な危機としての災害や戦争や大旱魃、あるいはゆっくりとした危機としての、たとえば、限界集落の出現や人口減少から生まれる問題群の中で、社会の中の何を重視するべきかの指針や判断を与えてくれるのではないか、といったことが挙げられる。
具体的に関連する現象として想定できるとして現象としては、環境/生態系破壊ないし保全や異常気象、生物多様性、絶滅危惧、生涯、一生、生死、生命倫理、限界集落、人口減少、等が挙げられるが、どのような現象に目を向けることになるのか、どのような現象が中心的な課題になっていくのかは、始めてみなければわからないし、それでよいと思っている。
本課題は一連の共同研究において通奏低音としてあった「環境」と、そこでの「生存」を意識化し、その極限的な局面を詳らかにすることを通して、人類の社会性(sociality)の進化についての理論構築を目指す試みである。初年度である本年度は、それぞれ個別の発表をしつつ、3つの学問分野において、「生存」、「環境」、「極限」といった概念がどのように扱われてきたか、あるいは、扱われうるのかを明確化し、これをメンバー間で、共有することを第一の目的とする。そのうえで、「環境における生存」をキーテーマに、その様態の極限を見定めるための方法を検討したい。
一連の共同研究は当初から長期プロジェクトとして企画され、群居性の霊長類の一員としての人類が進化の過程で獲得してきた高度な「社会性(sociality)」を種の誕生における最重要の特質として着目しようとするものであった。「社会性」なるものは、精神や社会の進化と同様に、化石として形に残りにくいものであるため、ヒトの社会とヒト以外の現生霊長類の社会との比較・検討が不可欠であった。そのため、霊長類学者と人類学者との共同研究が強く求められ、両者の共同討議として進められてきた。
今期は上記一連の共同研究の総括をするにあたり、「生存・環境・極限」といったやや広いテーマを中心課題として設定した。これは、研究主題としては「生存」を主軸とし、「生存(「生きる」ということ)の環境」を、その「極限」条件ないし「極限」状態まで見据えるものとした。ここで言う環境とは、生態学的かつ社会的であるような環境を指すが、そうした環境に「生きる」ことのぎりぎりの条件ないし状態を探ることを念頭に、これまでと同じ3つの学問分野である、霊長類社会・生態学、生態人類学、社会文化人類学からこの課題に迫りたい。
これまでの一連の共同研究は、ヒトを含む霊長類が適応したり発達させたりしてきた自然的/社会的環境を「舞台」ないし前提として展開されてきたものであり、複数の同種個体がともに<生>をはぐくむ術、生態学の用語を使えば「生存戦略」だが、そうしたものを明らかにすること、そして、人類が高度に発達させてきた「社会性」(sociality)の起原と進化の機構について考察することを目指してきた。
生物としての<生>(「生存」と言っても「生命」、「いのち」と言ってもよいが)、すなわち「生きる」ということは、まずは生物学的/生態学的に論じられうるものだが、それに一元的に還元することはむずかしい。とりわけわれわれ人類を含む霊長類が群居性動物として生きる存在である以上、そこには社会的なるものが必然的な要素としてあり、他者と「ともに生きて」いかねばならないということ、そしてそれ故に、その方法(生存戦略)は、非決定論的で、フレキシブル(可変的)で、コミュニケーショナルなものになると思われる。こうした性格については、すでに「集団」、「制度」、「他者」というこれまでの共同研究においても議論され、明らかにされてきたことでもあるが、この点に議論を焦点化して、人類の社会性の進化の解明に向けて新たな視座を追究していきたい。
本研究課題ではこれまでの一連の共同研究において明らかになっていたことを基盤として、その総括として、ヒトとヒト以外の霊長類の生物学的、生態学的環境の極限、および社会性の極限を詳らかにしていくことを通じて、人類社会における<生>=「生きること」の進化史的基盤を考えていくことを目指す。
以上のように、環境の極限を明らかにするとどのような学問的貢献があるかということの、より積極的な意義についても触れておきたい。まず、進化を遂げてきた社会の各段階のすべてに常に存在する「基盤」なるものが果たして何であるのかを明らかすることから、それは、「社会論」一般に寄与することになるだろうということが挙げられる。同時にそれは、社会のさまざまな危機、たとえば、急激な危機としての災害や戦争や大旱魃、あるいはゆっくりとした危機としての、たとえば、限界集落の出現や人口減少から生まれる問題群の中で、社会の中の何を重視するべきかの指針や判断を与えてくれるのではないか、といったことが挙げられる。
具体的に関連する現象として想定できるとして現象としては、環境/生態系破壊ないし保全や異常気象、生物多様性、絶滅危惧、生涯、一生、生死、生命倫理、限界集落、人口減少、等が挙げられるが、どのような現象に目を向けることになるのか、どのような現象が中心的な課題になっていくのかは、始めてみなければわからないし、それでよいと思っている。
本課題は一連の共同研究において通奏低音としてあった「環境」と、そこでの「生存」を意識化し、その極限的な局面を詳らかにすることを通して、人類の社会性(sociality)の進化についての理論構築を目指す試みである。初年度である本年度は、それぞれ個別の発表をしつつ、3つの学問分野において、「生存」、「環境」、「極限」といった概念がどのように扱われてきたか、あるいは、扱われうるのかを明確化し、これをメンバー間で、共有することを第一の目的とする。そのうえで、「環境における生存」をキーテーマに、その様態の極限を見定めるための方法を検討したい。
2. 少し長めの自己紹介
省略
3.1. 日常が崩壊するとき: 霊長類学から「生存の環境の極限」を考えるためのメモ(竹ノ下祐二)
この研究会は、「生存」を主軸にして、「生存の環境」をその「極限」条件まで見据え、生態学的・社会的環境に生きることのぎりぎりの条件と状態を探ることを念頭に、霊長類社会/生態学、生態人類学、社会文化人類学の3分野から統合的に迫ることを目的としている、らしい。 「生存の環境の極限」とは何だろうか。
「生存」を個体や社会集団、あるいは種が生きのびることと捉えると、生存の終焉とは、個体の死、集団の消滅/崩壊、個体群の絶滅、種の絶滅などである。煩雑になるのでこれらをひっくるめて「死」と呼ぼう。「生存の環境の極限条件」とは、この一線を超えると死が訪れる、というような境界のことだと考えればよいのだろうか。
だが、「死」はいっけん「生」の対義語のように思われるが、「生」の一部、「生」に織りこまれている事象と捉えることもできる。ニホンザルやチンパンジーの単位集団は世代を重ねて存続するが、そのした時間・空間スケールで眺めると、個体の死は日常のひとコマである。同様に、集団の消滅は地域個体群の歴史の中で、地域個体群の絶滅は種の歴史の中で、種の絶滅は系統の進化史の中で、ありふれた日常の「生」の一部となる。こう考えると、人類社会の進化史的基盤を探る本研究会で、単に「死」を「生」の彼岸に位置付け、その境界を「極限」とするのは具合が悪い。
非ヒト霊長類における「生存の環境の極限」を考える糸口として、本発表では「危機」について考える。霊長類学からみる危機をテーマとした、日本文化人類学会課題研究懇談会「危機の克服と地域コミュニティ」第9回研究会(2015年2月開催)において、五百部裕はマハレ山塊におけるアカコロブスを題材に議論を展開し、非ヒト霊長類には「危険」はあっても「危機」は存在しないのではないかと考察した [1]。マハレのアカコロブスはチンパンジーに捕食される。五百部によると、かれらは実際にチンパンジーに捕食されそうになると逃げたり抵抗したりするものの、チンパンジーに対して特段の対捕食者戦略を発達させているわけではないという。つまりアカコロブスにとって、チンパンジーによる捕食は日常に織り込みずみの事象であって、コミュニティレベルで「このままでは滅びてしまう」という危機感が共有されてはいないというのだ。そして、一般に非ヒト霊長類がそうした「危機感の共有」をおこなうかどうかは疑わしい。
コメンテーターであった私は「危険」と「危機」の違いを次のように考察した。危険 (danger) は「いま・ここ」に実在するが、危機 (threat) は想像力の産物である。危機とは、まず「いま・ここ」の状況から未来における滅亡を予測し、そのうえであらためて「いま・ここ」を未来と関連づけ「いま・ここ」のわれわれが、その線を越えたら不可避的に滅亡に向う境界線 (threshold) 上に位置すると再解釈したときに感じられるものである。このとき、境界線は実在ではなく、あくまで現実の解釈にともない創り出された想像の産物である。
そのように想像された境界線を「生存の環境の極限」と言いかえることが妥当なのかどうか、私にはよくわからないが、ここではとりあえずそう考えて先へ進むことにする。非ヒト霊長類は境界線を想像し創造することがあるのだろうか。あるとしたら、どのような状況だろうか。
物言わぬかれらの想像力は行動から推測するしかない。非ヒト霊長類が現在を「日常」の浮き沈みのひとコマではなく境界線を越えんとしている状況だと解釈したならば、かれらは日常的な対応を停止し、あらたな戦略を採用するはずである。したがって、非ヒト霊長類が危機的状況=極限状況にあるかどうかは、観察者からみてかれらが「死」に近いところにいるだけでは不十分で、その際にかれらが日常とは異なる振舞いをしているかどうかを検討する必要がある。そのような事例を4つ紹介したい。
一つ目の事例は、屋久島における群れの消滅と融合 である[2, 3]。屋久島のニホンザルは群れ密度が高く、群れの消滅がしばしば観察される。その際、すべてのオスを失ない最後に残されたメスが、隣接する群れに平和的に加入することがある。ニホンザル社会では、群れを移籍するのは通常オスのみである。
二つ目は、カフジ・ビエガ国立公園のヒガシローランドゴリラでみられた、複数メスの合同移籍である [4]。1990年代後半、内戦によって多くの核オスが殺害された際、メスたちが核オス抜きでまとまりを維持し、そろって他の集団に移籍したり、ヒトリゴリラを迎え入れて単雄群を再構築した。ゴリラでは、メスは個別に核オスとの関係を構築しており、通常メスたちが連れだって集団を移籍することはまれである。
三つ目は、嵐山のニホンザルにおける、アルファオスの交代にともなうアルファメスの行動変化である [5]。たてつづけにアルファオスが交代した群れで、通常は限られた血縁個体とした毛づくろいをしないアルファメスが、多くの非血縁メスと毛づくろいを行なうことで、結果として群れのまとまりを維持する役割を果たした。
四つ目は、東山動物園で、人口哺育されたゴリラのアカンボウの再導入プロセスにおいて見られた、シルバーバックとアカンボウの抱擁である。母親による後ろだてのないアカンボウと同居をはじめたシルバーバックが、ある日おもむろにアカンボウをしっかり抱擁したのである。
これらはいずれも、集団が崩壊したり秩序が乱れた際に、通常の社会構造・社会関係からは見られないような社会交渉が発現し、それによってなんとか集団が維持され、日常をとりもどした事例であると理解することができる。すなわち、日常が崩壊した際に非日常的な振舞いが発生し、それによって日常に回帰したといえる。特筆すべきは、こうした非日常的な振舞いが個体レベルではなく社会的反応として見られたことである。ここから、非ヒト霊長類においても、日常の崩壊すなわち危機的状況の認識と、その認識に関して社会の中にある種の「了解」が成立している可能性が示唆される。
References
[1] 五百部裕. 殺されることは『危機』なのか?―アカコロブスの対チンパンジー戦略―. 日本文化人類学会課題研究懇談会「危機の克服と地域コミュニティ」第9回研究会(2014年第3回研究会)2015年2月12日, 名古屋大学文学研究科, 名古屋市.
[2] Yukio Takahata, Shigeru Suzuki, Naobi Okayasu, and David Hill. Troop extinction and fusion in wild japanese macaques of yakushima island, japan. American Journal of Prima- tology, Vol. 33, No. 4, pp. 317–322, 1994.
[3] Hideki Sugiura, Naoki Agetsuma, and Shigeru Suzuki. Troop extinction and female fu- sion in wild japanese macaques in yakushima. International Journal of Primatology, Vol. 23, No. 1, pp. 69–84, 2002.
[4] Juichi Yamagiwa, John Kahekwa, and Augustin Kanyunyi Basabose. Infanticide and so- cial flexibility in the genus Gorilla. Primates, Vol. 50, No. 4, pp. 293–303, October 2009.
[5] 竹ノ下祐二. 政権交替と α メス 群れの統合維持に関わる α メスの役割. モンキー, 250–251, pp. 5–11, 1993.
「生存」を個体や社会集団、あるいは種が生きのびることと捉えると、生存の終焉とは、個体の死、集団の消滅/崩壊、個体群の絶滅、種の絶滅などである。煩雑になるのでこれらをひっくるめて「死」と呼ぼう。「生存の環境の極限条件」とは、この一線を超えると死が訪れる、というような境界のことだと考えればよいのだろうか。
だが、「死」はいっけん「生」の対義語のように思われるが、「生」の一部、「生」に織りこまれている事象と捉えることもできる。ニホンザルやチンパンジーの単位集団は世代を重ねて存続するが、そのした時間・空間スケールで眺めると、個体の死は日常のひとコマである。同様に、集団の消滅は地域個体群の歴史の中で、地域個体群の絶滅は種の歴史の中で、種の絶滅は系統の進化史の中で、ありふれた日常の「生」の一部となる。こう考えると、人類社会の進化史的基盤を探る本研究会で、単に「死」を「生」の彼岸に位置付け、その境界を「極限」とするのは具合が悪い。
非ヒト霊長類における「生存の環境の極限」を考える糸口として、本発表では「危機」について考える。霊長類学からみる危機をテーマとした、日本文化人類学会課題研究懇談会「危機の克服と地域コミュニティ」第9回研究会(2015年2月開催)において、五百部裕はマハレ山塊におけるアカコロブスを題材に議論を展開し、非ヒト霊長類には「危険」はあっても「危機」は存在しないのではないかと考察した [1]。マハレのアカコロブスはチンパンジーに捕食される。五百部によると、かれらは実際にチンパンジーに捕食されそうになると逃げたり抵抗したりするものの、チンパンジーに対して特段の対捕食者戦略を発達させているわけではないという。つまりアカコロブスにとって、チンパンジーによる捕食は日常に織り込みずみの事象であって、コミュニティレベルで「このままでは滅びてしまう」という危機感が共有されてはいないというのだ。そして、一般に非ヒト霊長類がそうした「危機感の共有」をおこなうかどうかは疑わしい。
コメンテーターであった私は「危険」と「危機」の違いを次のように考察した。危険 (danger) は「いま・ここ」に実在するが、危機 (threat) は想像力の産物である。危機とは、まず「いま・ここ」の状況から未来における滅亡を予測し、そのうえであらためて「いま・ここ」を未来と関連づけ「いま・ここ」のわれわれが、その線を越えたら不可避的に滅亡に向う境界線 (threshold) 上に位置すると再解釈したときに感じられるものである。このとき、境界線は実在ではなく、あくまで現実の解釈にともない創り出された想像の産物である。
そのように想像された境界線を「生存の環境の極限」と言いかえることが妥当なのかどうか、私にはよくわからないが、ここではとりあえずそう考えて先へ進むことにする。非ヒト霊長類は境界線を想像し創造することがあるのだろうか。あるとしたら、どのような状況だろうか。
物言わぬかれらの想像力は行動から推測するしかない。非ヒト霊長類が現在を「日常」の浮き沈みのひとコマではなく境界線を越えんとしている状況だと解釈したならば、かれらは日常的な対応を停止し、あらたな戦略を採用するはずである。したがって、非ヒト霊長類が危機的状況=極限状況にあるかどうかは、観察者からみてかれらが「死」に近いところにいるだけでは不十分で、その際にかれらが日常とは異なる振舞いをしているかどうかを検討する必要がある。そのような事例を4つ紹介したい。
一つ目の事例は、屋久島における群れの消滅と融合 である[2, 3]。屋久島のニホンザルは群れ密度が高く、群れの消滅がしばしば観察される。その際、すべてのオスを失ない最後に残されたメスが、隣接する群れに平和的に加入することがある。ニホンザル社会では、群れを移籍するのは通常オスのみである。
二つ目は、カフジ・ビエガ国立公園のヒガシローランドゴリラでみられた、複数メスの合同移籍である [4]。1990年代後半、内戦によって多くの核オスが殺害された際、メスたちが核オス抜きでまとまりを維持し、そろって他の集団に移籍したり、ヒトリゴリラを迎え入れて単雄群を再構築した。ゴリラでは、メスは個別に核オスとの関係を構築しており、通常メスたちが連れだって集団を移籍することはまれである。
三つ目は、嵐山のニホンザルにおける、アルファオスの交代にともなうアルファメスの行動変化である [5]。たてつづけにアルファオスが交代した群れで、通常は限られた血縁個体とした毛づくろいをしないアルファメスが、多くの非血縁メスと毛づくろいを行なうことで、結果として群れのまとまりを維持する役割を果たした。
四つ目は、東山動物園で、人口哺育されたゴリラのアカンボウの再導入プロセスにおいて見られた、シルバーバックとアカンボウの抱擁である。母親による後ろだてのないアカンボウと同居をはじめたシルバーバックが、ある日おもむろにアカンボウをしっかり抱擁したのである。
これらはいずれも、集団が崩壊したり秩序が乱れた際に、通常の社会構造・社会関係からは見られないような社会交渉が発現し、それによってなんとか集団が維持され、日常をとりもどした事例であると理解することができる。すなわち、日常が崩壊した際に非日常的な振舞いが発生し、それによって日常に回帰したといえる。特筆すべきは、こうした非日常的な振舞いが個体レベルではなく社会的反応として見られたことである。ここから、非ヒト霊長類においても、日常の崩壊すなわち危機的状況の認識と、その認識に関して社会の中にある種の「了解」が成立している可能性が示唆される。
References
[1] 五百部裕. 殺されることは『危機』なのか?―アカコロブスの対チンパンジー戦略―. 日本文化人類学会課題研究懇談会「危機の克服と地域コミュニティ」第9回研究会(2014年第3回研究会)2015年2月12日, 名古屋大学文学研究科, 名古屋市.
[2] Yukio Takahata, Shigeru Suzuki, Naobi Okayasu, and David Hill. Troop extinction and fusion in wild japanese macaques of yakushima island, japan. American Journal of Prima- tology, Vol. 33, No. 4, pp. 317–322, 1994.
[3] Hideki Sugiura, Naoki Agetsuma, and Shigeru Suzuki. Troop extinction and female fu- sion in wild japanese macaques in yakushima. International Journal of Primatology, Vol. 23, No. 1, pp. 69–84, 2002.
[4] Juichi Yamagiwa, John Kahekwa, and Augustin Kanyunyi Basabose. Infanticide and so- cial flexibility in the genus Gorilla. Primates, Vol. 50, No. 4, pp. 293–303, October 2009.
[5] 竹ノ下祐二. 政権交替と α メス 群れの統合維持に関わる α メスの役割. モンキー, 250–251, pp. 5–11, 1993.
3.2. 「生存・環境・極限」:生態人類学から考えるための話題集(杉山祐子)
1. はじめに
ヒトはその歴史のなかで、移動をくりかえして分布域を拡げ、集団の規模や構成のバリエーションも拡大してきた。そこには利用できる環境の拡大や食性の変化、人口構造の変化などを伴い、ある場合にははからずも、ある場合には意識的な「技術革新」を生みながら現在の姿にたどりついたとみることができる。
ヒトと環境の相互作用の系を生計活動の詳細な検討から明らかにしてきた生態人類学の立場から考えるとき、「生存」は、時間幅および「集団」の単位や消費・再生産の単位を多層的にとらえることに関わってくる。
2. 生計活動と環境
生態人類学初期の研究は、現代の狩猟採集民が資源を使い尽くすことはなく、むしろ余裕をもって生活していることを明らかにした(田中1971、丹野1986)。それは長年にわたる知識や技術の蓄積があるから可能だともいえる。しかし狩りや採集活動で、「効率」が悪くなるとさっさと移動するというやりかたは(市川1996)環境利用の基本形と考えてよいだろう。
ヒトと自然が相互に馴化しあう関係も特徴的である。熱帯雨林では狩猟採集民の伐木などの人間活動が植物の多様性を生み出している(市川1996,2010)。ミオンボ林帯では、火入れなどの人間活動が火に強い木々を育て、それらが作る環境が焼畑農耕の基盤となっている。結果的にではあれ、人間活動と自然が相互依存的に独特の環境をつくりだしてきたことがわかる。より積極的に自然に働きかけ、特定の循環を生み出すことによって成り立つ生業もある。マテンゴのンゴロ農法(加藤2002)、アリのエンセーテ(重田 1996)やバナナ栽培(丸尾 2002)など多くの在来農法はその例である。
さらに、現代のヒトの生業にきわだつ特徴は、異なる自然環境下で異なる生業を営む人びとが「共生」や交易などを通じて、たがいの環境を自身の生活圏としていることだろう。寺嶋(1997)は、エフェが森での狩猟採集、レッセが村での農耕という異なる生業を営みながらも相互に深い関わりをもつことを示して、「森も村も自分たちの世界と接するもうひとつの世界であり、つまりは自己の世界を拡大するもの(寺嶋 1997:204)」と述べる。人間がその分布域を広げる過程では、それぞれの環境に適応した生業とその技術を開発してきたと考えられるが、自分たちとは異なる環境に異なる生業をもって暮らす他集団との関係を築くことによって、人間はその生存の可能性を大きくひろげたといえよう。
3. 生存と「極限」を時間枠と「関係」からみる
上記のことをふまえると、「極限」をかんがえるとき、異なる時間枠で、集団の生成・成長のサイクルや環境との関係を考慮に入れる必要があることがわかる。それは長期的時間枠と中短期的時間枠の2種類である。
長期的時間枠とは10年〜30年以上で、世代交代を視野に入れた時間の長さを考える。ここでの生活の様態は移動と一定期間の定住の組み合わせによって成り立つ。移動と定住の組み合わせは、異なる自然環境利用の組み合わせとも対応しうるし、集団の規模や新しい集団の生成、他の集団との交易等を含めた相互扶助的関係の構築も関わる。次の3つの場合が考えられる。
a) 極相的「極限」:「集団」の発達段階における「極相」としての「極限」。人口規模や集団の構成が極相状態を示す。技術革新や新しい資源の開発などはあるだろうが、人口が一定以上に増加した場合、移動その他の方法による人口コントロールがおこなわれる。また、他の生態環境圏とのセットで生活圏が構成される場合も考えられる。
b) 展開の「極限」:この場合はさらに、2つのサブタイプが考えられる。
①集団サイクル・循環型:集団の組成が次のサイクルに移るときの「極限」。ベンバの村の例では、新しい村が創設されて発展するが、世代交代期にはもめごとが起きて分裂、その後、また新しい村が創られる循環をくりかえす。
②分蜂・フロンティア型:マテンゴなどの例がこれにあたる。中心村の人口密度が高まると、中心村とは異なる手法で未開拓地に新村を作り、フロンティアを開発しながら展開する。
c) 消滅の「極限」:生存のぎりぎり。干ばつや飢饉、疫病等による人口減少、他集団との抗争による集団の消滅などが考えられる。ただし、資源が少ないことが必ずしも人びとを競合に向かわせるわけではない。また、こうした消滅にむかう「極限」をきりぬける方途がフロンティアを開拓し、移動や食性の変化を伴いながら新たな適応の様態を生み出す可能性もある。
中短期的時間枠での「極限」を考えるには、消費の単位の幅(ひとり・母子・炉帯と世帯、「共食」を許す単位を日常的な文脈と飢饉などの危機的(極限)状況下の文脈でとらえることや、日常的にみられる小さな極限(日常における社会的葛藤、呪い、暴力など)を個人と中間的集まり、集団全体、および集団間関係など多層的に検討する必要があると思われる。
5. まとめ
以上、生態人類学から「生存・環境・極限」をかんがえるとき、1) ヒト=環境系を長期的なスケールでの移動・定住を含むダイナミズムとしてとらえる生態史的視点、2)日常における多様な消費の単位の組み合わせと生存の単位を多層的にとらえる視点、3)環境の大きな変化など現代の諸課題を組み込む視点などが重要である。
引用文献
市川光雄 1996「文化の変異と社会統合 ― ムブティ・ピグミーの植物利用にみられる変異」田中・ 掛谷・市川・太田編『続・自然社会の人類学 』アカデミア 出版会:409-437
------- 2010 「アフリカ熱帯雨林の歴史生態学に向けて」『森棲みの生態誌』京都大学出版会
重田眞義2002 「アフリカにおける持続的な集約農業の可能性」掛谷誠編『アフリカ農耕民の世界』京都大学出版会
田中二郎 1972 『ブッシュマン』思索社
寺嶋秀明 1997『共生の森』東京大学出版会
丸尾聡 2002 「バナナとともに生きる人びと―タンザニア北西部・ハヤの村から」掛谷誠編『アフリカ農耕民の世界―素の在来性と変容―』京都大学出版会
ヒトはその歴史のなかで、移動をくりかえして分布域を拡げ、集団の規模や構成のバリエーションも拡大してきた。そこには利用できる環境の拡大や食性の変化、人口構造の変化などを伴い、ある場合にははからずも、ある場合には意識的な「技術革新」を生みながら現在の姿にたどりついたとみることができる。
ヒトと環境の相互作用の系を生計活動の詳細な検討から明らかにしてきた生態人類学の立場から考えるとき、「生存」は、時間幅および「集団」の単位や消費・再生産の単位を多層的にとらえることに関わってくる。
2. 生計活動と環境
生態人類学初期の研究は、現代の狩猟採集民が資源を使い尽くすことはなく、むしろ余裕をもって生活していることを明らかにした(田中1971、丹野1986)。それは長年にわたる知識や技術の蓄積があるから可能だともいえる。しかし狩りや採集活動で、「効率」が悪くなるとさっさと移動するというやりかたは(市川1996)環境利用の基本形と考えてよいだろう。
ヒトと自然が相互に馴化しあう関係も特徴的である。熱帯雨林では狩猟採集民の伐木などの人間活動が植物の多様性を生み出している(市川1996,2010)。ミオンボ林帯では、火入れなどの人間活動が火に強い木々を育て、それらが作る環境が焼畑農耕の基盤となっている。結果的にではあれ、人間活動と自然が相互依存的に独特の環境をつくりだしてきたことがわかる。より積極的に自然に働きかけ、特定の循環を生み出すことによって成り立つ生業もある。マテンゴのンゴロ農法(加藤2002)、アリのエンセーテ(重田 1996)やバナナ栽培(丸尾 2002)など多くの在来農法はその例である。
さらに、現代のヒトの生業にきわだつ特徴は、異なる自然環境下で異なる生業を営む人びとが「共生」や交易などを通じて、たがいの環境を自身の生活圏としていることだろう。寺嶋(1997)は、エフェが森での狩猟採集、レッセが村での農耕という異なる生業を営みながらも相互に深い関わりをもつことを示して、「森も村も自分たちの世界と接するもうひとつの世界であり、つまりは自己の世界を拡大するもの(寺嶋 1997:204)」と述べる。人間がその分布域を広げる過程では、それぞれの環境に適応した生業とその技術を開発してきたと考えられるが、自分たちとは異なる環境に異なる生業をもって暮らす他集団との関係を築くことによって、人間はその生存の可能性を大きくひろげたといえよう。
3. 生存と「極限」を時間枠と「関係」からみる
上記のことをふまえると、「極限」をかんがえるとき、異なる時間枠で、集団の生成・成長のサイクルや環境との関係を考慮に入れる必要があることがわかる。それは長期的時間枠と中短期的時間枠の2種類である。
長期的時間枠とは10年〜30年以上で、世代交代を視野に入れた時間の長さを考える。ここでの生活の様態は移動と一定期間の定住の組み合わせによって成り立つ。移動と定住の組み合わせは、異なる自然環境利用の組み合わせとも対応しうるし、集団の規模や新しい集団の生成、他の集団との交易等を含めた相互扶助的関係の構築も関わる。次の3つの場合が考えられる。
a) 極相的「極限」:「集団」の発達段階における「極相」としての「極限」。人口規模や集団の構成が極相状態を示す。技術革新や新しい資源の開発などはあるだろうが、人口が一定以上に増加した場合、移動その他の方法による人口コントロールがおこなわれる。また、他の生態環境圏とのセットで生活圏が構成される場合も考えられる。
b) 展開の「極限」:この場合はさらに、2つのサブタイプが考えられる。
①集団サイクル・循環型:集団の組成が次のサイクルに移るときの「極限」。ベンバの村の例では、新しい村が創設されて発展するが、世代交代期にはもめごとが起きて分裂、その後、また新しい村が創られる循環をくりかえす。
②分蜂・フロンティア型:マテンゴなどの例がこれにあたる。中心村の人口密度が高まると、中心村とは異なる手法で未開拓地に新村を作り、フロンティアを開発しながら展開する。
c) 消滅の「極限」:生存のぎりぎり。干ばつや飢饉、疫病等による人口減少、他集団との抗争による集団の消滅などが考えられる。ただし、資源が少ないことが必ずしも人びとを競合に向かわせるわけではない。また、こうした消滅にむかう「極限」をきりぬける方途がフロンティアを開拓し、移動や食性の変化を伴いながら新たな適応の様態を生み出す可能性もある。
中短期的時間枠での「極限」を考えるには、消費の単位の幅(ひとり・母子・炉帯と世帯、「共食」を許す単位を日常的な文脈と飢饉などの危機的(極限)状況下の文脈でとらえることや、日常的にみられる小さな極限(日常における社会的葛藤、呪い、暴力など)を個人と中間的集まり、集団全体、および集団間関係など多層的に検討する必要があると思われる。
5. まとめ
以上、生態人類学から「生存・環境・極限」をかんがえるとき、1) ヒト=環境系を長期的なスケールでの移動・定住を含むダイナミズムとしてとらえる生態史的視点、2)日常における多様な消費の単位の組み合わせと生存の単位を多層的にとらえる視点、3)環境の大きな変化など現代の諸課題を組み込む視点などが重要である。
引用文献
市川光雄 1996「文化の変異と社会統合 ― ムブティ・ピグミーの植物利用にみられる変異」田中・ 掛谷・市川・太田編『続・自然社会の人類学 』アカデミア 出版会:409-437
------- 2010 「アフリカ熱帯雨林の歴史生態学に向けて」『森棲みの生態誌』京都大学出版会
重田眞義2002 「アフリカにおける持続的な集約農業の可能性」掛谷誠編『アフリカ農耕民の世界』京都大学出版会
田中二郎 1972 『ブッシュマン』思索社
寺嶋秀明 1997『共生の森』東京大学出版会
丸尾聡 2002 「バナナとともに生きる人びと―タンザニア北西部・ハヤの村から」掛谷誠編『アフリカ農耕民の世界―素の在来性と変容―』京都大学出版会
3.3. 「社会文化人類学における『生存』」(春日直樹)
社会文化人類学の歴史を3期に分けることにより、各期で生存がどのように論じられてきたのかを編年的に総括して、今後の研究方向についての話題提供をおこなう。なお、ここでいう「生存」とは、極限的な状況を配慮しつつ生きるという意味である。
1.1870年代〜1910年代
社会文化人類学が学問として確立した時期であり、進化主義や伝搬主義などさまざまな立場から、人類の社会と文化を論じる研究が現れた。とくにイギリスでは、エドワード・タイラーの『原始文化』やジェームズ・フレーザーの『金枝篇』が、進化の観点から洞察を展開した。ともに壮大なスケールで人類史を振り返り、とりわけ人間の思考の進化を明らかにしようとしたが、そこでは「死」「屍体」「睡眠」「夢」「影」などが中心的な契機となって思考を活発化した、とされている。すべて人間の生存にかかわる重大事項であるがゆえに、人びとを捉えてはなさなかったわけである。
この時期の社会文化人類学を特色づけるのは、長期的な視野に立った分析であり、同世紀の他の諸分野にも容易にみいだすことができる。とくに経済学は長期に経済の成長を論じるがゆえに、生存かつかつの生活を将来の到達点とするような「陰気な科学」であった。『金枝篇』の最後は陰気とはいえないが、生存に対する不安を宇宙大の規模で描いており、同時代の知識人に多大な影響を及ぼした。
この時期の研究から学ぶものとは、数世紀・数十世紀あるいはさらに長期の視野から生存を論じる姿勢であり、さらにはタイラーの「文化」概念のような総合的な観点である。
2.1920年代〜1980年代
社会文化人類学は専門性を高めて、現地調査に力点を置いた詳細なデータ収集と緻密な分析が主流となった。調査期間の限定があり、かつ調査対象の歴史的資料の乏しさもあって、研究は総じて長期的な視野を失った。ちょうど経済学において1923年のケインズが長期的アプローチを批判し、「長期には[=結局のところ]われわれは皆死んでしまう」(『貨幣改革論』)として短期的アプローチを唱道した頃と重なるが、経済学がこれによって生存の危機に瀕した人びとを救おうとしたのに対して、社会文化人類学はむしろ、調査地の人びとの生活が生存維持のために営まれているのでなく、彼ら固有の価値や規範や動機などに基盤を置くことを主張するようになったので、生存というテーマへの直面を避ける傾向が生まれた。食糧不足を扱った民族誌も数は数少なく存在するのだが、一般的にいえば生存に言及しない記述スタイルが主流を占める。
生存の危機に瀕している状況は、むしろ植民地化や貨幣経済が引き起こす例外的なものとして扱われたといえる。この点は、マーシャル・サーリンズが提起した「最初の豊かな社会」の概念によく現れている。そこでは、採集狩猟民の労働時間の短さ、余暇の長さ、食糧に関する楽観的な姿勢を強調する一方で、食糧事情を心配する採集狩猟民については言及しないまま、ただ現在の彼らが辺境に追いやられた特異な状況にある点を指摘している。
3.1980年代後半〜現在
第二期で生存の問題にふれた数少ない民族誌は、その状況で文化がどのように変容し、あるいは再構築されるかに主眼を置いていた。これに対して近年では、「生存を文化的状況として考察すること」「生存としての文化」をめぐる研究が着実に現れつつある、ということができる。それらを大きく分類すれば、「プロジェクト」研究と「生社会」研究になろう。
「プロジェクト」研究とは、災害、気候変動、生物多様性などに関する大型プロジェクトの設計・実践・評価のプロセスを対象にして、大なり小なり科学技術論とくにアクターネットワーク理論(ANT)の影響を受けている。生存の危機とかかわるプロジェクトではあるが、結局は何らかの「発展」を目指しているので、生存の問題から遠ざかるようになってしまう(ただし、難民キャンプを対象にしたプロジェクト研究については、こうした特徴づけはできず、全般的にオーソドックスな民族誌的な形態をとることが多い)。
「生社会」の研究の具体的なテーマは、不適切な診断と治療、特定の疾病のスティグマ化、際限のない延命措置、過剰な薬物投与、ローカルな病気の放置、伝統的な療法の抑圧と消失、臓器売買、信仰に反した屍体処理、など実に多岐にわたる。不安や危険にさらされる身体や人格がどのようにして生まれるか、そしてこの苦境の中でいかに主体としての自己を再構築するかに主眼が置かれている。ただしポール・ラビノーなどの一部をのぞけば、「生社会」「生権力」あるいは「むき出しの生」といった流行の概念を使用することを避ける傾向にある。こうした表現自体がすでに特定のイメージを喚起する意味で文化である、という認識をもつからであろう。
以上のように社会文化人類学の研究を編年的に振り返れば、生存の問題を文化の観点から研究すべし、という結論を導けそうな気がするが、このまま受け入れるのは抵抗がある。文化と自然という二項対比自体が、「科学」対「人文学」という西洋近代の知識に関する二分類に由来している。これに対するラディカルな批判として、近年は「多自然主義」を提起する人類学者もでているが、残念ながら説得力に乏しい。
とりあえずは、社会文化人類学の慣れ親しんだ民族誌において、生存を直接にテーマ化しないが生存の問題と不可分である記述があちこちに散りばめられていることに、注意を喚起したい。人類学者が人びとの生活を総体的に理解しようと努めるかぎり、そこには生きること、死ぬことにかかわる事象が当然ながら観察できる。生存の問題は、宗教、呪術、親族などの諸領域に分かれてすでに民族誌的に記述されている。これらの記述から生存の問題を掘り起こし、たとえば「生社会」「むき出しの生」がテーマ化される状況における生存と比較する作業は、それなりに有意義であろう。
1.1870年代〜1910年代
社会文化人類学が学問として確立した時期であり、進化主義や伝搬主義などさまざまな立場から、人類の社会と文化を論じる研究が現れた。とくにイギリスでは、エドワード・タイラーの『原始文化』やジェームズ・フレーザーの『金枝篇』が、進化の観点から洞察を展開した。ともに壮大なスケールで人類史を振り返り、とりわけ人間の思考の進化を明らかにしようとしたが、そこでは「死」「屍体」「睡眠」「夢」「影」などが中心的な契機となって思考を活発化した、とされている。すべて人間の生存にかかわる重大事項であるがゆえに、人びとを捉えてはなさなかったわけである。
この時期の社会文化人類学を特色づけるのは、長期的な視野に立った分析であり、同世紀の他の諸分野にも容易にみいだすことができる。とくに経済学は長期に経済の成長を論じるがゆえに、生存かつかつの生活を将来の到達点とするような「陰気な科学」であった。『金枝篇』の最後は陰気とはいえないが、生存に対する不安を宇宙大の規模で描いており、同時代の知識人に多大な影響を及ぼした。
この時期の研究から学ぶものとは、数世紀・数十世紀あるいはさらに長期の視野から生存を論じる姿勢であり、さらにはタイラーの「文化」概念のような総合的な観点である。
2.1920年代〜1980年代
社会文化人類学は専門性を高めて、現地調査に力点を置いた詳細なデータ収集と緻密な分析が主流となった。調査期間の限定があり、かつ調査対象の歴史的資料の乏しさもあって、研究は総じて長期的な視野を失った。ちょうど経済学において1923年のケインズが長期的アプローチを批判し、「長期には[=結局のところ]われわれは皆死んでしまう」(『貨幣改革論』)として短期的アプローチを唱道した頃と重なるが、経済学がこれによって生存の危機に瀕した人びとを救おうとしたのに対して、社会文化人類学はむしろ、調査地の人びとの生活が生存維持のために営まれているのでなく、彼ら固有の価値や規範や動機などに基盤を置くことを主張するようになったので、生存というテーマへの直面を避ける傾向が生まれた。食糧不足を扱った民族誌も数は数少なく存在するのだが、一般的にいえば生存に言及しない記述スタイルが主流を占める。
生存の危機に瀕している状況は、むしろ植民地化や貨幣経済が引き起こす例外的なものとして扱われたといえる。この点は、マーシャル・サーリンズが提起した「最初の豊かな社会」の概念によく現れている。そこでは、採集狩猟民の労働時間の短さ、余暇の長さ、食糧に関する楽観的な姿勢を強調する一方で、食糧事情を心配する採集狩猟民については言及しないまま、ただ現在の彼らが辺境に追いやられた特異な状況にある点を指摘している。
3.1980年代後半〜現在
第二期で生存の問題にふれた数少ない民族誌は、その状況で文化がどのように変容し、あるいは再構築されるかに主眼を置いていた。これに対して近年では、「生存を文化的状況として考察すること」「生存としての文化」をめぐる研究が着実に現れつつある、ということができる。それらを大きく分類すれば、「プロジェクト」研究と「生社会」研究になろう。
「プロジェクト」研究とは、災害、気候変動、生物多様性などに関する大型プロジェクトの設計・実践・評価のプロセスを対象にして、大なり小なり科学技術論とくにアクターネットワーク理論(ANT)の影響を受けている。生存の危機とかかわるプロジェクトではあるが、結局は何らかの「発展」を目指しているので、生存の問題から遠ざかるようになってしまう(ただし、難民キャンプを対象にしたプロジェクト研究については、こうした特徴づけはできず、全般的にオーソドックスな民族誌的な形態をとることが多い)。
「生社会」の研究の具体的なテーマは、不適切な診断と治療、特定の疾病のスティグマ化、際限のない延命措置、過剰な薬物投与、ローカルな病気の放置、伝統的な療法の抑圧と消失、臓器売買、信仰に反した屍体処理、など実に多岐にわたる。不安や危険にさらされる身体や人格がどのようにして生まれるか、そしてこの苦境の中でいかに主体としての自己を再構築するかに主眼が置かれている。ただしポール・ラビノーなどの一部をのぞけば、「生社会」「生権力」あるいは「むき出しの生」といった流行の概念を使用することを避ける傾向にある。こうした表現自体がすでに特定のイメージを喚起する意味で文化である、という認識をもつからであろう。
以上のように社会文化人類学の研究を編年的に振り返れば、生存の問題を文化の観点から研究すべし、という結論を導けそうな気がするが、このまま受け入れるのは抵抗がある。文化と自然という二項対比自体が、「科学」対「人文学」という西洋近代の知識に関する二分類に由来している。これに対するラディカルな批判として、近年は「多自然主義」を提起する人類学者もでているが、残念ながら説得力に乏しい。
とりあえずは、社会文化人類学の慣れ親しんだ民族誌において、生存を直接にテーマ化しないが生存の問題と不可分である記述があちこちに散りばめられていることに、注意を喚起したい。人類学者が人びとの生活を総体的に理解しようと努めるかぎり、そこには生きること、死ぬことにかかわる事象が当然ながら観察できる。生存の問題は、宗教、呪術、親族などの諸領域に分かれてすでに民族誌的に記述されている。これらの記述から生存の問題を掘り起こし、たとえば「生社会」「むき出しの生」がテーマ化される状況における生存と比較する作業は、それなりに有意義であろう。
第2回研究会での発表内容
1. 生きる共同体にとっての過去と未来──死者・神・自然(内堀基光)
人間の社会性(人と人との関係のしかた)というときの「人間らしさ」は、過去との結びつき(被規定性、制度の惰性性、記憶)と未来への視点(配慮、不安、希望、存続、そして極限的なものとしての恐怖)にある。すべてこれが出発点である。現在は過去と未来のバランスの軸心だが、過去の重みと未来の重みは、人間の集合としての共同体によって大きく異なる。ここで共同体はごく小さい家族的単位から、最大限では全人類にまでおよぶものと考えることにする。個体としての人間は、二者を超える複数のとき、記憶と期待を直接には共有しない。これを共有するものを共同体と考えるが、共有の程度はさまざまであり、それが共同体というかたちで現れた人間の社会性の具体的性質と限界を決める。全人類の集合の中で、さまざまの共同体が空間的に併存し、また重層的に積み重なっている。
上の段落の後半は人間の社会性≈共同性について当たり前のことであって、人間にかかわるかぎり、その集団論、制度論、他者論のすべての前提になっていたことである。その延長に位置する「生存・極限」研究では、制度研論文・他者研論文で書ききれなかったこと、つまり前段落前半分を論じることにする。とりあえず、制度研・他者研論文の到着点からはじめることにしたいと考えている。それは人間に特有の二重世界の成立に関わることであり、およそ次のような言い方になる。
「死者がかつて生きていた、おそらくは多くの場合空間的、心理的に近いところにいた(親しかったか憎んでいたか)他個体との個体間関係に基礎を置くのに対し、はるかに無限定で、共同体の了解、承認によってのみ同定が可能となるような幻影としての他者(精霊や神)の場合、前者(死者)の存在には必要でない世界の認識が必要になる。おそらくは「もう一つの世界」としての空間、世界の二重性。死者の存在と他界の存在を単純には同一の初発事象として結びつけられなかったのだが、幻影としての他者の空間が用意されているかぎり、死者の居場所もまた用意されたというべきであろう。本質的にエフェメラルでもありうる死者に対して、精霊は、確固とした共同了解の意味世界に、幻影でありながら幻影を超えたものとして置かれている。その世界は尋常の人間にとってはある種外部世界であり、精霊はそこからより身近な覚醒時の生活世界に去来してくる。このもう一つの世界の成立によって、死者も精霊と同じ資格で独立した存在者になりうるのである。」
(人類の)歴史は、個体の死があるから、生成すると言ったのはヘーゲルだが、20世紀後半は広い意味で、さまざまな意味での「反歴史」の時代だったように思われる。21世紀はふたたび(良くも悪くも)歴史の時代になるのかもしれない。歴史はとりあえず過去に起きたこととして語られるが、過去の語りには未来の不安、恐怖の投影があり、歴史はいわばそれを視ようとする共同性をめざした営為となる。今ここにあるものと、今ここにないもの、今ここにあって、いつかなくなる(かもしれない)ものについての、不安、恐怖をともなう想いは、アニミズムとは相対立するものでもあろう。そこにはアニミズムの二重世界とは異なる、未来と予測、また恐怖と救済というモメントが働くことになる。「神」のイメージを導入しても良い。アニミズムは永遠の現在を拠り所とするが、単独者としての個体の「神」の永遠性は、虚偽の永続、虚偽の過去と未来、つまりフィクションとしての永遠の謂である。「神」は、永遠としての、掴みえない未来における恐怖と救済のバランシングのなかに出現する。あるいはまたアニミズムでもありえた「死者と過去」から、恐怖を内在した「神と未来」への転換をそこに見ることもできる。
ここから共同体の2つの対照的な存続根拠についての議論を展開することがこの研究プロジェクトにおける私の課題となる。予告としては、共同体の過去と現在から未来への永続(持続)への根拠を語る議論として、内山節『共同体の基礎理論』(2010)、Peter Metcalfのロングハウス・コミュニティ史(Life History of Longhouse,2009)を読み解くことから始めるつもりである。あわせて、岩田慶治によるアニミズム論の再評価も課題として設定する。
上の段落の後半は人間の社会性≈共同性について当たり前のことであって、人間にかかわるかぎり、その集団論、制度論、他者論のすべての前提になっていたことである。その延長に位置する「生存・極限」研究では、制度研論文・他者研論文で書ききれなかったこと、つまり前段落前半分を論じることにする。とりあえず、制度研・他者研論文の到着点からはじめることにしたいと考えている。それは人間に特有の二重世界の成立に関わることであり、およそ次のような言い方になる。
「死者がかつて生きていた、おそらくは多くの場合空間的、心理的に近いところにいた(親しかったか憎んでいたか)他個体との個体間関係に基礎を置くのに対し、はるかに無限定で、共同体の了解、承認によってのみ同定が可能となるような幻影としての他者(精霊や神)の場合、前者(死者)の存在には必要でない世界の認識が必要になる。おそらくは「もう一つの世界」としての空間、世界の二重性。死者の存在と他界の存在を単純には同一の初発事象として結びつけられなかったのだが、幻影としての他者の空間が用意されているかぎり、死者の居場所もまた用意されたというべきであろう。本質的にエフェメラルでもありうる死者に対して、精霊は、確固とした共同了解の意味世界に、幻影でありながら幻影を超えたものとして置かれている。その世界は尋常の人間にとってはある種外部世界であり、精霊はそこからより身近な覚醒時の生活世界に去来してくる。このもう一つの世界の成立によって、死者も精霊と同じ資格で独立した存在者になりうるのである。」
(人類の)歴史は、個体の死があるから、生成すると言ったのはヘーゲルだが、20世紀後半は広い意味で、さまざまな意味での「反歴史」の時代だったように思われる。21世紀はふたたび(良くも悪くも)歴史の時代になるのかもしれない。歴史はとりあえず過去に起きたこととして語られるが、過去の語りには未来の不安、恐怖の投影があり、歴史はいわばそれを視ようとする共同性をめざした営為となる。今ここにあるものと、今ここにないもの、今ここにあって、いつかなくなる(かもしれない)ものについての、不安、恐怖をともなう想いは、アニミズムとは相対立するものでもあろう。そこにはアニミズムの二重世界とは異なる、未来と予測、また恐怖と救済というモメントが働くことになる。「神」のイメージを導入しても良い。アニミズムは永遠の現在を拠り所とするが、単独者としての個体の「神」の永遠性は、虚偽の永続、虚偽の過去と未来、つまりフィクションとしての永遠の謂である。「神」は、永遠としての、掴みえない未来における恐怖と救済のバランシングのなかに出現する。あるいはまたアニミズムでもありえた「死者と過去」から、恐怖を内在した「神と未来」への転換をそこに見ることもできる。
ここから共同体の2つの対照的な存続根拠についての議論を展開することがこの研究プロジェクトにおける私の課題となる。予告としては、共同体の過去と現在から未来への永続(持続)への根拠を語る議論として、内山節『共同体の基礎理論』(2010)、Peter Metcalfのロングハウス・コミュニティ史(Life History of Longhouse,2009)を読み解くことから始めるつもりである。あわせて、岩田慶治によるアニミズム論の再評価も課題として設定する。
2. 社会の進化と種間関係(足立薫)
動物の社会行動を扱う生物学の分野にあって、異種間の関係は中心的な課題となることは非常に稀であった。その理由の一つは、進化生物学が同種個体群の中の個体変異に注目するために、種間関係は二次的なテーマとならざるを得ないという点にある。とくに近年の進化生物学では、個体変異や進化過程の遺伝的基盤への関心が強化され、社会行動の進化は主流の課題と言えなくなりつつある。加えて、「種とは何か」を定義するのが困難であるという、古典的な問題の存在が、種間の社会関係をとらえることを一層難しくしている。ローレンツやティンバーゲンが動物の社会行動を研究し始めたエソロジーの黎明期においては、種特異的な定型的行動が、動物のコミュニケーション研究の主要な対象であった。そのため、種は均一な性質をもった対象と捉えられ、種間の特徴を比較して進化が議論された。それに対して社会行動の研究に行動生態学の概念がもたらされて以降は、進化適応の単位が種ではなく個体、あるいは遺伝子に変更されたことによって、同種個体間の行動の変異を明らかにすることが、社会進化研究の重要な課題となり、種間の違いへの関心は薄れていくことになった。
社会進化の行動学的な研究では、種という単位が主題化されなくなる一方で、生態系の群集構造を対象とする進化生態学では、種間関係は一貫して重要な分野を構成してきた。進化生態学では種間関係を、共進化の文脈で考察する。被食―捕食や寄生―宿主関係は、その代表例であり、また種間での利益の偏りが極端なものでもある。同所的に共存する異種個体群の関係は、生物多様性や生態系のバランス、安定性の側面から、環境に関わる重要な現代的課題として取り扱われている。
霊長類学においては、社会集団における個体の社会行動を対象とする分野では、種間関係は主要な研究対象とはなってこなかった。その例外が、混群研究である。混群は異種の群れが同所的に生息するだけでなく、ともに採食や遊動を行い、同種の群れのように生活する現象である。熱帯地域の中型小型の霊長類にみられる現象で、ほかに鳥類や魚類でも観察されている。霊長類の混群研究は、群集生態学における競争と共存の理論を基礎として発展してきた。近縁な異種の群れの個体どうしが、潜在的な競合をどのように回避しているのか、あるいは競合のコストを上回る利益をどのように得ているのかについて、個体の行動から混群形成の要因を探る試みが行われてきた。ここでは混群を形成するという社会行動における種間関係は、競合にかかわるコストとベネフィットの収支という生態学的仮説に置き換えられている。
一方で、混群は、異種の群れが同所的に共存しながらゆるやかに群れとしてのまとまりを保ち続ける現象である。混群形成の現象では、同種群のようなグルーミングや順位を争う闘争行動のような、目立った社会交渉は見られないが、「そこはかとない関心」を保ちながら異種の個体どうしがひとつの群れにとどまり続ける。ゆるやかに結びつきながら、出会ったり離れたりを繰り返す混群では、どのようなメカニズムで集団が形成されているのか、同種群と同様のアプローチでその社会性に迫ることは難しい。目立った社会交渉ではなく、従来は社会行動とは考えられなかった遊動や採食行動の詳細なレベルでの同調や共同の様子を、時空間に再現するような新たな分析手法が必要とされている。このような混群独自の社会のありように注目し、異種間の社会関係の進化を構想することの利点は何だろうか。
種間関係に注目をして混群社会を見る場合、「種」の違いに注目することによって、個体間の差異が相対的に捨象され、遺伝的変異に視野を限定せずに行動を見ることができるだろう。さらに「群集」のレベルに視線をおくことによって、群れ全体や個体群全体という大きなレベルで社会をとらえ、個体の意図や意識を前提にせずに行動を社会性の文脈におくことが可能になる。なぜ群れを作るのか、という問いから動物の社会行動に迫ろうとするならば、本当に明らかにしたいのは「群居性の構え」とか「同調への志向性」といった基本的な特性である。これらの基本特性は説明されるべき社会性に対する前提条件ではなく、まさに社会性そのものであると理解でき、そのような理解によってはじめて人間も含めた動物の社会性の進化へのアプローチが有意味なものになると考えられる。
混群における種間関係から社会行動を見ることで、「構え」や「同調」を前提としない社会性を構想できるとしたが、事態は同種個体間の関係でも同様である。実体的な「関係」に基づく因果関係ではなく、どんな枠組みも先行しないもっとも基本的なところで、集団が成り立つ様子をとらえることが重要となる。一つの方法としてルーマンの二重の偶有性による社会システムの創発を参照し、「相手が自分が予想することとは違う行為を行うかもしれない」というダブル・コンティンジェンシーから社会性の成り立ちを明らかにすることが考えられる。「解決」をゴールに志向するのではなく、「失敗」をニュートラルに組み込んだ社会システムとして社会をとらえることができれば、混群のような特異な社会集団も含めて、多元的な社会のありようを統一的に進化の枠組みで扱うことが可能になるだろう。
社会進化の行動学的な研究では、種という単位が主題化されなくなる一方で、生態系の群集構造を対象とする進化生態学では、種間関係は一貫して重要な分野を構成してきた。進化生態学では種間関係を、共進化の文脈で考察する。被食―捕食や寄生―宿主関係は、その代表例であり、また種間での利益の偏りが極端なものでもある。同所的に共存する異種個体群の関係は、生物多様性や生態系のバランス、安定性の側面から、環境に関わる重要な現代的課題として取り扱われている。
霊長類学においては、社会集団における個体の社会行動を対象とする分野では、種間関係は主要な研究対象とはなってこなかった。その例外が、混群研究である。混群は異種の群れが同所的に生息するだけでなく、ともに採食や遊動を行い、同種の群れのように生活する現象である。熱帯地域の中型小型の霊長類にみられる現象で、ほかに鳥類や魚類でも観察されている。霊長類の混群研究は、群集生態学における競争と共存の理論を基礎として発展してきた。近縁な異種の群れの個体どうしが、潜在的な競合をどのように回避しているのか、あるいは競合のコストを上回る利益をどのように得ているのかについて、個体の行動から混群形成の要因を探る試みが行われてきた。ここでは混群を形成するという社会行動における種間関係は、競合にかかわるコストとベネフィットの収支という生態学的仮説に置き換えられている。
一方で、混群は、異種の群れが同所的に共存しながらゆるやかに群れとしてのまとまりを保ち続ける現象である。混群形成の現象では、同種群のようなグルーミングや順位を争う闘争行動のような、目立った社会交渉は見られないが、「そこはかとない関心」を保ちながら異種の個体どうしがひとつの群れにとどまり続ける。ゆるやかに結びつきながら、出会ったり離れたりを繰り返す混群では、どのようなメカニズムで集団が形成されているのか、同種群と同様のアプローチでその社会性に迫ることは難しい。目立った社会交渉ではなく、従来は社会行動とは考えられなかった遊動や採食行動の詳細なレベルでの同調や共同の様子を、時空間に再現するような新たな分析手法が必要とされている。このような混群独自の社会のありように注目し、異種間の社会関係の進化を構想することの利点は何だろうか。
種間関係に注目をして混群社会を見る場合、「種」の違いに注目することによって、個体間の差異が相対的に捨象され、遺伝的変異に視野を限定せずに行動を見ることができるだろう。さらに「群集」のレベルに視線をおくことによって、群れ全体や個体群全体という大きなレベルで社会をとらえ、個体の意図や意識を前提にせずに行動を社会性の文脈におくことが可能になる。なぜ群れを作るのか、という問いから動物の社会行動に迫ろうとするならば、本当に明らかにしたいのは「群居性の構え」とか「同調への志向性」といった基本的な特性である。これらの基本特性は説明されるべき社会性に対する前提条件ではなく、まさに社会性そのものであると理解でき、そのような理解によってはじめて人間も含めた動物の社会性の進化へのアプローチが有意味なものになると考えられる。
混群における種間関係から社会行動を見ることで、「構え」や「同調」を前提としない社会性を構想できるとしたが、事態は同種個体間の関係でも同様である。実体的な「関係」に基づく因果関係ではなく、どんな枠組みも先行しないもっとも基本的なところで、集団が成り立つ様子をとらえることが重要となる。一つの方法としてルーマンの二重の偶有性による社会システムの創発を参照し、「相手が自分が予想することとは違う行為を行うかもしれない」というダブル・コンティンジェンシーから社会性の成り立ちを明らかにすることが考えられる。「解決」をゴールに志向するのではなく、「失敗」をニュートラルに組み込んだ社会システムとして社会をとらえることができれば、混群のような特異な社会集団も含めて、多元的な社会のありようを統一的に進化の枠組みで扱うことが可能になるだろう。
3. 暴力と平和力:ソロモン諸島の「民族紛争」渦中における生存戦略(藤井真一)
本報告の着想は二つある。ひとつは西田正規[2003; 2004]による武力と平和力の共進化に関する議論である。彼は、直立二足歩行に伴う手の自由使用から、武器へと転じうる道具の携帯へと進化する中で、人類は強力な武力を手に入れたという議論を展開した。さらに、強力な武力を社会内部へ向けることは社会や種の存続を危機に晒すとの推論から、「より強い武力が進化するには、その武力が種内にむかって暴発することを避けるための、より強力な平和力の進化がともなっていなくてはならない」[西田 2004: 276]と論じた。
もうひとつは、メラネシア人類学の主要な研究主題である戦争論と交換論である。ニューギニアの部族間抗争やソロモン諸島の首狩りに関する事例報告は膨大にある。わずかだが、暴力的な事態を収拾するための方策に関する報告もある[Wedgwood 1930; Epstein (ed.) 1974など]。特に、報復殺人(blood feud)を終結させるための儀礼において贈与されるモノが、紛争解決のみならず婚資としても機能して、敵対関係を婚姻同盟へと転換させるとの議論は注目に値する[Harrison 1993; Knauft 1999など]。
本報告では、西田による武力と平和力の共進化に関する議論と、メラネシア人類学における戦争と交換に関する議論に着想を得て、報告者が調査しているソロモン諸島の「民族紛争」を題材に、暴力と平和力の相互的関係について考えることを目指した。
1998年末から2003年にかけてソロモン諸島で生じた「民族紛争」はガダルカナル島民とマライタ島出身者の対立だとされてきた。しかし、「民族」を対立軸に、社会・経済・政治的背景を無視した紛争理解では不十分との指摘が繰り返されており、紛争の背景には植民地主義の遺産、経済や土地をめぐる諸問題などの複合的要因があったといわれる。死者数は約200名、国内避難者数は35,000名と推計されており、従来の研究では避難者数の約4割を占めるマライタ島出身者に対する注目が多かった。一方、同じく約4割近いガダルカナル島内の避難民についての報告は数少ない。本報告では、紛争渦中に避難生活を強いられたガダルカナル島民、特に激戦地であったガダルカナル島北東部の人びとの生存戦略に焦点を当てて取り上げた。
ガダルカナル島北東部の人びとは約6割が住居を失い、ブッシュへ逃げるしかなかったといわれる。当該地域には大規模なアブラヤシ農園が位置し、特にマライタ島からの労働者とその親族が多く居住していた。「民族紛争」勃発を受けて彼らのほとんどが住居や財産を捨てて首都への避難やマライタ島への帰還を余儀なくされた。また、ガダルカナル島出身者たちも、紛争に巻き込まれることを回避すべく、自身の畑に仮住まいを建てつつ複数地点に点在する畑を転々として生き延びた。
武装集団へ積極的に加担した情報提供者によれば、畑仕事などのため出払っているマライタ系住民の家屋に対して、農薬等散布用のポンプで石油を撒いて放火して回ることで「居場所」を失わせたという。その他、ガダルカナル島北東部の人びとによる紛争への消極的な関与として、海へ潜って太平洋戦争時の弾薬を収集したり、海岸部でマライタ側武装集団が襲撃してこないかどうか警備したりしていたことが挙げられる。
一方、紛争への加担や関与を回避する消極的対応として、次のような事例があった。(1)加入を要請する武装集団に対して食糧等を提供することにより戦闘員の提供を回避すること。(2)ガダルカナル島北東部の人間と通婚して当該地域に居住していた一部のマライタ系住民を暴力から守るため、「ペル・ウル」と呼ばれる贈与財の授受を伴った儀礼を行なわせることで紛争から距離をとらせたこと。(3)近隣にあるゲラ島との混血であることを強みに、封鎖された陸路ではなく海路で生活物資を調達してきてガダルカナル島北東部の集落で分配したことなどである。
これら紛争を回避する消極的対応はいずれも、贈与が暴力の発現を未然に防ぐ平和力を発揮する事例と考えることができる。ただし、それらはポトラッチのような競争的な性格ではなく、受贈者に負債を与えるようなものでもない、モノを贈ることで非物質的な社会関係を享受する「異次元交換」であった。そのような平和力を発揮する贈与を可能としたのは、豊かな食糧をもたらすガダルカナル島北東部の生態環境でもあった。
こうした贈与、交換、分配を通じた敵対的なかかわりの回避は類人猿にも認められ、進化史的基盤を考える上で恰好の素材を提供するはずである。ただし、交換(贈与)されたモノにそれまでの社会関係の来歴が織り込まれる人類の交換は、食物を独占したいという我欲を克服して分配する類人猿の行為と区別できそうである。ここにヒトとサルの進化の連続性と非連続性、暴力と平和力の共進化の筋道を見出せるかもしれない。
■参照文献
もうひとつは、メラネシア人類学の主要な研究主題である戦争論と交換論である。ニューギニアの部族間抗争やソロモン諸島の首狩りに関する事例報告は膨大にある。わずかだが、暴力的な事態を収拾するための方策に関する報告もある[Wedgwood 1930; Epstein (ed.) 1974など]。特に、報復殺人(blood feud)を終結させるための儀礼において贈与されるモノが、紛争解決のみならず婚資としても機能して、敵対関係を婚姻同盟へと転換させるとの議論は注目に値する[Harrison 1993; Knauft 1999など]。
本報告では、西田による武力と平和力の共進化に関する議論と、メラネシア人類学における戦争と交換に関する議論に着想を得て、報告者が調査しているソロモン諸島の「民族紛争」を題材に、暴力と平和力の相互的関係について考えることを目指した。
1998年末から2003年にかけてソロモン諸島で生じた「民族紛争」はガダルカナル島民とマライタ島出身者の対立だとされてきた。しかし、「民族」を対立軸に、社会・経済・政治的背景を無視した紛争理解では不十分との指摘が繰り返されており、紛争の背景には植民地主義の遺産、経済や土地をめぐる諸問題などの複合的要因があったといわれる。死者数は約200名、国内避難者数は35,000名と推計されており、従来の研究では避難者数の約4割を占めるマライタ島出身者に対する注目が多かった。一方、同じく約4割近いガダルカナル島内の避難民についての報告は数少ない。本報告では、紛争渦中に避難生活を強いられたガダルカナル島民、特に激戦地であったガダルカナル島北東部の人びとの生存戦略に焦点を当てて取り上げた。
ガダルカナル島北東部の人びとは約6割が住居を失い、ブッシュへ逃げるしかなかったといわれる。当該地域には大規模なアブラヤシ農園が位置し、特にマライタ島からの労働者とその親族が多く居住していた。「民族紛争」勃発を受けて彼らのほとんどが住居や財産を捨てて首都への避難やマライタ島への帰還を余儀なくされた。また、ガダルカナル島出身者たちも、紛争に巻き込まれることを回避すべく、自身の畑に仮住まいを建てつつ複数地点に点在する畑を転々として生き延びた。
武装集団へ積極的に加担した情報提供者によれば、畑仕事などのため出払っているマライタ系住民の家屋に対して、農薬等散布用のポンプで石油を撒いて放火して回ることで「居場所」を失わせたという。その他、ガダルカナル島北東部の人びとによる紛争への消極的な関与として、海へ潜って太平洋戦争時の弾薬を収集したり、海岸部でマライタ側武装集団が襲撃してこないかどうか警備したりしていたことが挙げられる。
一方、紛争への加担や関与を回避する消極的対応として、次のような事例があった。(1)加入を要請する武装集団に対して食糧等を提供することにより戦闘員の提供を回避すること。(2)ガダルカナル島北東部の人間と通婚して当該地域に居住していた一部のマライタ系住民を暴力から守るため、「ペル・ウル」と呼ばれる贈与財の授受を伴った儀礼を行なわせることで紛争から距離をとらせたこと。(3)近隣にあるゲラ島との混血であることを強みに、封鎖された陸路ではなく海路で生活物資を調達してきてガダルカナル島北東部の集落で分配したことなどである。
これら紛争を回避する消極的対応はいずれも、贈与が暴力の発現を未然に防ぐ平和力を発揮する事例と考えることができる。ただし、それらはポトラッチのような競争的な性格ではなく、受贈者に負債を与えるようなものでもない、モノを贈ることで非物質的な社会関係を享受する「異次元交換」であった。そのような平和力を発揮する贈与を可能としたのは、豊かな食糧をもたらすガダルカナル島北東部の生態環境でもあった。
こうした贈与、交換、分配を通じた敵対的なかかわりの回避は類人猿にも認められ、進化史的基盤を考える上で恰好の素材を提供するはずである。ただし、交換(贈与)されたモノにそれまでの社会関係の来歴が織り込まれる人類の交換は、食物を独占したいという我欲を克服して分配する類人猿の行為と区別できそうである。ここにヒトとサルの進化の連続性と非連続性、暴力と平和力の共進化の筋道を見出せるかもしれない。
■参照文献
- 西田正規 2003 「社会進化と平和力」西田正規ほか編『人間性の起源と進化』63-95頁、昭和堂。
- 西田正規 2004 「家族社会の進化と平和力―家族・分配・平等性」寺嶋秀明編『平等と不平等をめぐる人類学的研究』274-290頁、ナカニシヤ出版。
- Epstein, A.L. (ed.) 1974 Contention and Dispute: Aspects of Law and Social Control in Melanesia. Canberra: Australian National University Press.
- Harrison, Simon 1993 The Mask of War: Violence, Ritual and the Self in Melanesia. Manchester & New York: Manchester University Press.
- Knauft, Bruce M. 1999 From Primitive to Postcolonial in Melanesia and Anthropology. Ann Arbor: The University of Michigan Press.
- Wedgwood, Camilla H. 1930 Some Aspects of Warfare in Melanesia. Oceania 1(1): 5-33.
第3回研究会での発表内容
1.類人猿のホーティカルチャー(黒田末寿)
生きものと環境の関係は複雑である。ここではその関係のひとつを類人猿の採食活動が再植樹に及ぼす影響から考える。
1)動物が自己の外部をつくる
動物は存在することで自己の「外部」を作る。外部の例は自己の殻・巣・構築物、排泄物などがあり、それが他の生物や同種の生息環境を構成するという多様で親和的あるいは対立的な諸形態がある。
親和的な外部の形成は、A代謝の老廃物で身体を覆う(歯・角質皮膚・角の起源、サンゴ、貝など)、B外部材で身体を覆う(巣、蓑虫の殻、ビーバーのダム、衣服・住居など)がある。さらに、C環境利用が利用環境の安定化・増大をもたらすメカニズム(フジツボが岩を溶かして居場所を作るなど、原半栽培:動物と植物・バクテリアの双利的な関係)、D利用環境を保護する半栽培(Cを主体が意図しておこなう)、E利用生物の増大をはかる生殖管理行為(栽培)などがある。
ここで報告するのは、類人猿が採食行動によって環境をより利用価値を高くするCに分類できる現象である。Cでよく知られている現象は類人猿による果樹の種子散布(c1)であるが、ここではそれに加えて採食樹の変形(c2)について述べる。Cは通常、正のフィードバック構造をもつ。種子散布c1は採食・排出が環境を再生産する。採食樹の変形c2は枝折り(剪定)破壊が利用環境の維持に貢献する。前者は個体と環境の相互作用が結果的に種の環境改善をもたらすのに対し、後者は個体の行為が同種あるいは他種の採食を助ける社会的性格をもつ。
2)種子散布
果実をつける樹種の多くは、動物に食べられ種子を親木から離れた場所に排泄される
=散布されることによって繁殖する。なかでも固い殻や大きな種子をもつ果実は鳥や小型の霊長類が食べて運ぶことができないので、もっぱら類人猿やゾウの種子散布に頼ることになる。このような種子は、消化管(酸と温熱)を通ることで休眠が解け発芽する性質を持つ。また、消化によって発芽を阻害する種子を覆う果肉を取り除く。
一方、動物による咀嚼と消化は種子にダメージを与えるので、それに対処する方法として果実はつぎのような方略をもつ。①大きな固い殻の種子、②小さい無数の種子、③甘いゼリーと苦い有毒な種。これらパターンは中間的なものもあり、また特定の動物との関係とリンクする。
3)種子散布者としての類人猿とゾウ
ゾウはとくに①の果樹の種子散布に重要。
糞による種子散布は覆いとスカラベが種子を地下に引き込む作用で齧歯類による食害を防ぐが、ゾウのように糞が多すぎるとカビとキノコでダメージを受けることがある。ゴリラ程度の糞が最適。ボノボとチンパンジーの糞からの発芽はゴリラに比べてなぜか劣る(定量分析が必要)が、数が多いので問題にはならない。類人猿は木の実の果実を食べて種子を散布し、オプティマルハビタットを再生産しており、果樹の方もそれに適応している。
4)ゴリラの半栽培
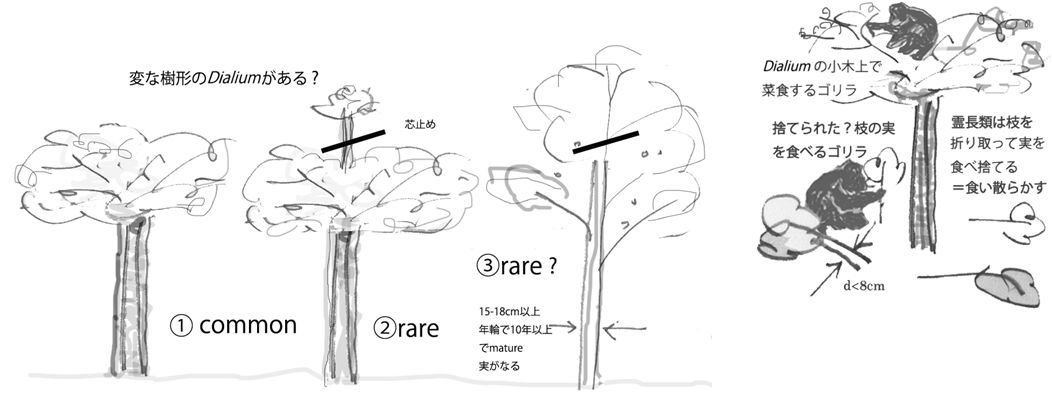
コンゴ共和国のヌアバレ・ドキ国立公園(以後ドキ)ではツエゴチンパンジーとニシテイチゴリラが共存する。ドキでは低木のカキであるDiospiros sp.と高木Dialium sp. はいずれも優占種である。低木のカキは樹高10m以下の細長い木であるが、ほぼ80%以上が根元から折り曲げられ地上近くに横ばいになっている。その状況やゴリラの採食行動の観察からこれはゴリラが変形したと考えられる。まっすぐに伸びたカキは、幹が細すぎておとなのゴリラには昇れないが、このようにして誰もが菜食できる状態になっている。
Dialium sp.は30m以上になる大木で、10年で10mほどになり結実し始める。サルやチンパンジーも食べるが、ゴリラの糞からの発芽が一番多い。この木には高さ10mほどで真ん中の枝が折られ(芯止め)、枝が横に広がっている変形された木がよく見られる。折りとられた枝の太さから変形樹はゴリラの仕業と推測されるが、どこまで意識して樹形を変形しているのかは不明である。
Dialiumは細長く伸びても、大木になってもツルがない限りゴリラには昇れないが、高さ10m–15mでとどまる限り、そばの小木から移ることが可能なので、幹を芯止めして高木にさせなければ、体格が大きいおとなのゴリラも恒久的に利用できる。変形樹にはサルがあがっていてもチンパンジーがあがっているのを見たことがない。ゴリラ専用の樹としてチンパンジーは敬遠するのかもしれない。ゴリラが意識的にこうした変形をおこなっているかどうかを確認できていないので、この行為をDの「半栽培」と判定できないが、栽培の起源を考えるときに多くの示唆を与えてくれる。
1)動物が自己の外部をつくる
動物は存在することで自己の「外部」を作る。外部の例は自己の殻・巣・構築物、排泄物などがあり、それが他の生物や同種の生息環境を構成するという多様で親和的あるいは対立的な諸形態がある。
親和的な外部の形成は、A代謝の老廃物で身体を覆う(歯・角質皮膚・角の起源、サンゴ、貝など)、B外部材で身体を覆う(巣、蓑虫の殻、ビーバーのダム、衣服・住居など)がある。さらに、C環境利用が利用環境の安定化・増大をもたらすメカニズム(フジツボが岩を溶かして居場所を作るなど、原半栽培:動物と植物・バクテリアの双利的な関係)、D利用環境を保護する半栽培(Cを主体が意図しておこなう)、E利用生物の増大をはかる生殖管理行為(栽培)などがある。
ここで報告するのは、類人猿が採食行動によって環境をより利用価値を高くするCに分類できる現象である。Cでよく知られている現象は類人猿による果樹の種子散布(c1)であるが、ここではそれに加えて採食樹の変形(c2)について述べる。Cは通常、正のフィードバック構造をもつ。種子散布c1は採食・排出が環境を再生産する。採食樹の変形c2は枝折り(剪定)破壊が利用環境の維持に貢献する。前者は個体と環境の相互作用が結果的に種の環境改善をもたらすのに対し、後者は個体の行為が同種あるいは他種の採食を助ける社会的性格をもつ。
2)種子散布
果実をつける樹種の多くは、動物に食べられ種子を親木から離れた場所に排泄される
=散布されることによって繁殖する。なかでも固い殻や大きな種子をもつ果実は鳥や小型の霊長類が食べて運ぶことができないので、もっぱら類人猿やゾウの種子散布に頼ることになる。このような種子は、消化管(酸と温熱)を通ることで休眠が解け発芽する性質を持つ。また、消化によって発芽を阻害する種子を覆う果肉を取り除く。
一方、動物による咀嚼と消化は種子にダメージを与えるので、それに対処する方法として果実はつぎのような方略をもつ。①大きな固い殻の種子、②小さい無数の種子、③甘いゼリーと苦い有毒な種。これらパターンは中間的なものもあり、また特定の動物との関係とリンクする。
3)種子散布者としての類人猿とゾウ
ゾウはとくに①の果樹の種子散布に重要。
糞による種子散布は覆いとスカラベが種子を地下に引き込む作用で齧歯類による食害を防ぐが、ゾウのように糞が多すぎるとカビとキノコでダメージを受けることがある。ゴリラ程度の糞が最適。ボノボとチンパンジーの糞からの発芽はゴリラに比べてなぜか劣る(定量分析が必要)が、数が多いので問題にはならない。類人猿は木の実の果実を食べて種子を散布し、オプティマルハビタットを再生産しており、果樹の方もそれに適応している。
4)ゴリラの半栽培
コンゴ共和国のヌアバレ・ドキ国立公園(以後ドキ)ではツエゴチンパンジーとニシテイチゴリラが共存する。ドキでは低木のカキであるDiospiros sp.と高木Dialium sp. はいずれも優占種である。低木のカキは樹高10m以下の細長い木であるが、ほぼ80%以上が根元から折り曲げられ地上近くに横ばいになっている。その状況やゴリラの採食行動の観察からこれはゴリラが変形したと考えられる。まっすぐに伸びたカキは、幹が細すぎておとなのゴリラには昇れないが、このようにして誰もが菜食できる状態になっている。
Dialium sp.は30m以上になる大木で、10年で10mほどになり結実し始める。サルやチンパンジーも食べるが、ゴリラの糞からの発芽が一番多い。この木には高さ10mほどで真ん中の枝が折られ(芯止め)、枝が横に広がっている変形された木がよく見られる。折りとられた枝の太さから変形樹はゴリラの仕業と推測されるが、どこまで意識して樹形を変形しているのかは不明である。
Dialiumは細長く伸びても、大木になってもツルがない限りゴリラには昇れないが、高さ10m–15mでとどまる限り、そばの小木から移ることが可能なので、幹を芯止めして高木にさせなければ、体格が大きいおとなのゴリラも恒久的に利用できる。変形樹にはサルがあがっていてもチンパンジーがあがっているのを見たことがない。ゴリラ専用の樹としてチンパンジーは敬遠するのかもしれない。ゴリラが意識的にこうした変形をおこなっているかどうかを確認できていないので、この行為をDの「半栽培」と判定できないが、栽培の起源を考えるときに多くの示唆を与えてくれる。
2.極限的出会いと進化:ヒトは何とどのように出会い,どう進化してきたのか?(寺嶋秀明)
ヒトは進化史的に見れば,その誕生以来,さまざまな<異なるもの>との出会いを経て現在に至る。ヒトがヒト以外のものと出会ったときにどのようなことが起こり,その後,両者の間にはどのような関係が成立し,どのような結果をもたらしたのか考えてみたい。<異なるもの>とは,生命をもたない物質の場合もあれば,自分以外の別種のヒトの場合もある。片方が消滅したという大事件も何度か起こっている。<異なるもの>との出会いとは,後から省みると,その瞬間からヒトあるいはヒトが出会ったものの生活や運命が大きく変わるという特異点(シンギュラリティ)でもある。そういった出会いを<極限的な出会い>としていくつか考えてみたい。
1)ヒトと<石>
チンパンジーの雄がディスプレイのために大きな石を投げたり,また雌が堅いナッツを割るために自然石をハンマーとして使うという報告がある(Boesch 1991)。ニホンザルでも石を用いた一人遊びがある(ハフマン 1991)。初期の人類もなんらかの形で自然石をそのまま用いていたことは十分想像できる。そのようなヒトと石との関係は,約250万年前,現生人類へとつながる初期人類の仲間であるホモ・ハビリスが,石と石をぶつけ合って一方の石を割った時点から大きく変わった。一方の石にできた鋭利な破断面が動物や植物などを切るための刃物としてきわめて有用であることがわかったのである。オルドワン・タイプと呼ばれる石器の誕生である。進化史的にはチンパンジーとヒトが別れたのがおよそ700万年前といわれているので,人類誕生から450万年ほど後の出来事であった。
上記の出来事は,その後のヒトの運命に大きな影響を与えたことは疑いがないが,石にとってはどうだろうか。石は本来ヒトとなんらの関係がなくても存在する。しかし「石器」という事物の登場によって,ヒトにとっては石はたんなる自然物ではなく,道具性を内包した素材となった。石からすると,ヒトは素材性を現実化するエージェントになったのである。石器はホモ・ハビリスのオルドワン・タイプからホモ・エレクトゥスのアシューレアン,ネアンデルタールのムステリンアンを経て,サピエンスのオーリニャシアン石器文化へと進化した。新しい文化では,それまで潜在的であった素材性が,新しい形態の石器として現実に利用されるようになる。これをヒト(の加工技術)と石(の素材性)の共進化と考えてみたい。
2)ヒトと野生動物・植物
ヒトと自然界の動物・植物との関係はたいへん重要であるが,食物源としての動植物との出会いはことのほか大きな意味をもつ。ヒトは生存のための食物を自然界に求めるが,それらは生得的に決まっているものではない。さまざまな動植物の中から,ヒトはその時その時の状況に合わせて,これといったものを発見し,食物レパートリーに加えていく。食物レパートリーとして選ばれた動植物とヒトの間には新しい関係が成立する。後期旧石器時代初頭の人類は,それ以前と比べかなり広範に,貝類や魚類など水棲の動物の利用やウサギなどショウほ乳類の狩猟を開始したことが知られている (Kuhn and Steiner 2006)。たぶん,最終氷期のきびしい環境条件への対応であろうが,この食物レパートリーの拡大がヒトの学習能力と環境適応力を増大し,ユーラシアの広大な大地に分布を広げる基盤になったことは十分推測しうる。食物レパートリーの広さは,現生の多くの狩猟採集民においても大きな特徴となっている。
はじめは互いに自然界の風景の一要素に過ぎなかったもの同士が,ヒトが利用することにより特別な関係に入る。ヒトはそれら動植物をより効率的にできるようさまざまな工夫を重ねていく。一方,動植物はなるべくヒトからの被害を避けて生きるようになるが,むしろその繁殖の一端を捕食者に委ねる傾向を持つもの出現した。その極限的形態が野生動物の家畜化や野生植物の栽培作物化である。ヒトと動植物の一部は太いボンドを形成し共に進化して今日にいたっている。ヒトとそれらの動植物は,どちらももはや単独では存続不可能である。これについては,後に改めて論じたい。
3)サピエンスとネアンデルタール
サピエンスはおよそ10万年前,誕生の地アフリカを出立し,5万年ほど前にレバント地方を経由して,ヨーロッパに入ってきたといわる。しかし,そこにはすでにサピエンスのいわば従兄弟であるネアンデルタールが25万年以上の長きにわたって住んでいた。そして両者はおよそ45,000年から40,000年前の間,ヨーロッパの地で共存することになった。両者の出会いのようすは確たる証拠もなく謎に包まれているが,武力衝突などの形跡は見つかっていないようだ。さらに近年はネアンデルタールの人骨を試料としたゲノム解析が進み,アフリカ大陸以外の現生人類には2〜5%ほどのネアンデルタールの遺伝子が存在することが明らかになった。両者は出会ったのみならず,交配し,混血するまでになったのであった(ペーボ 2015)。
ネアンデルタールの絶滅の原因については,いろいろと論争がある。現在のところ,最終氷河期の強烈な気候変動(ハインリヒイベント5)のために,ネアンデルタールはヨーロッパの南の温暖地方への後退による分布域減少ならびに人口減少に直面していたところへ,サピエンスという同じニッチを求める競争者の進出も重なって,良好なニッチ占拠が思うようにいかず,また骨太で筋肉量多しという高エネルギー消費体格ゆえに,それまで大きく依存してきた大型の獲物の減少によって,栄養学的に問題が生じ,急激に個体数も減らしたのであろうと考えられている。交替劇初期にはネアンデルタールとサピエンスの文化程度はそう違わなかったという(佐野・大森 2015)。しかしサピエンスは,新環境への進出過程において急速に学習能力を発達させ,生業の技術やシステムの改良,および社会的リフォームをおこない,ネアンデルタールでは不可能であった激しく変動する環境への適応にも成功しえたと想像されている (佐野・大森 2015)。
ところで,それぞれの出会いの地でサピエンスはネアンデルタールとどのように交流し,別れたのであろうか。近年の研究によるとネアンデルタールは昔想像されたような獣じみた野蛮人ではなく,むしろサピエンスにとって魅力的であったかもしれない。両者の間には文化交流や愛の交換も確実にあった。その結果サピエンスはネアンデルタールが長い間のヨーロッパでの適応過程で獲得したと思われる白い肌や青い瞳(いずれも弱い太陽光への適応),そばかすといった形質や,ある種の免疫機構を獲得したといわれる。また,同時に,花粉症やアレルギー,うつ,喫煙癖などの現代人を悩ませている病気などもネアンデルタール由来かもしれないという(Nature vol. 523: 512-513, 2015)。
サピエンスとネアンデルタール人との出会いは,結果的には,生き残ったサピエンスが一方的にうまくやったことのように見られているが,ネアンデルタールの存在の影響も否定できない。ある割合の遺伝子がしっかりと残っていることを考えるならば,勝った敗けたという観点から両者の出会いを見ることは短慮に過ぎる。
4)サピエンスと動物,その新しい出会い
ヒトと野生の動植物との出会いについてはすでにふれたが,交替劇の前後におけるサピエンスと動物との関係の進化は象徴的意味を含むという点で,一段階レベルを異にした「新しい出会い」と呼ぶべきものと考えられる。サピエンスの現代的行動の一つが象徴的意味を含む行動であるが,それはアフリカの中石器時代にすでに生まれている (McBreaty and Brooks 2000) 。たとえば,南アフリカ・ブロンボス洞窟の7〜9万年前の地層からは,幾何学的文様が線刻されたオーカー塊,貝に穴を開け紐でつないだネックレスなどが発見されている。それらはあきらかにたんなる実用を越えたものとして制作されたに違いない。ただし,ヒトの進化の上では画期的出来事であるが,アフリカにおけるそれらの出現は時間・空間的に散発的であり,同一文化内でそういった行動がそのまま発達したという形跡はない(門脇 2013)。
ところがヨーロッパにおける交替劇が完了した4万年前以降,フランコ=カンタブリア地域の洞窟絵画や南ドイツの小彫像文化など,動物をモチーフにした象徴的表現が急速に南西ヨーロッパからドイツにかけて広がった。ショーベやアルタミラ,ラスコーなどの洞窟絵画でわかるように,それらはとても「原始美術」として絵画技術の発展段階を示すようなものではなく,現代の審美的水準でも際だった芸術性を示している。ある時点から野生動物がヒトの象徴的活動の焦点になった理由としては,狩猟対象としての獲物を捕るための呪術説や儀礼説などいろいろな解釈が提出されているが,そこにはたんなる呪術的手段を越えた動物の姿を認める必要がある。ヒトと動物との関係性,あるいは,ヒトが動物を見るその見方の大きな変化である。
G・バタイユは『ラスコー,あるいは芸術の誕生』 (Bataille 1955) において「人間性」の発見と「動物性」の発見を同一視している。ヒトは自己についての意識の高まりとともに人間性を発見するのと同時に動物の動物性を発見した。躍動美,生命力,神秘性,自由などによって表現される動物の動物性とは,もともと動物であったはずの人間がその人間性の発達と引き替えに失った聖なる特性であるとされる。洞窟絵画のモチーフである動物は「食物としての動物」ではなく,ヒトが失った聖なる特性の具現化,ヒトよりも神に近い存在としてのイメージであり,見えない心的存在の可視化と解釈すべきである。ほとんどの洞窟壁画がなぜ,ヒトが容易に近づけないような洞窟の奥,暗闇に閉じ込められた場所にばかり描かれているのかという疑問への答えの一つは,表象されるべき動物たちには,ヒトが想像力で補完してはじめてその全体を現前しうるような描かれ方,鑑賞法が必要であったということかもしれない。
そのような人間と動物との分離は,一方で,両者の間に捕食者と獲物との関係などの自然的関係を越えた相互関係を生み出すことになる。野生動物との「対話による交渉」は,現生人類,とくに自然と密接な関わりをもつ狩猟採集民などではごく普通におこなわているものだ。それによって両者の間には共感的なコミュニケーションの回路が開かれ,ヒトと野生動物とは互酬的関係に入る(寺嶋 2007, 大村 2007)。動物と共感的にコミュニケーションし,ともに今を生きる存在として両立できるという関係の発見は,はからずも,その後のサピエンスの文化に絶大な影響を及ぼすことになった。それがおおよそ3万年この方の,野生動物の家畜化,および野生植物の栽培植物化であった。ヒトと野生の動植物との共感的出会いが文明への第一歩を導いたのである。
5)サピエンスと人工知能
人類の進化を推進してきた「交替劇」はサピエンスで完成,終了したわけではないはずである。では,サピエンスの次に何が来るのか,しばしば議論の的になる。現在のサピエンスのように生活の大きな部分を「文化的環境」によって覆われている生物にとっては,これまでのような自然淘汰の原理が働かないともいわれる。たしかに,自然要因による淘汰圧はもう進化の原動力とはならないかもしれない。しかし,人為による文化環境の大きな改変があるとしたら,そのような改変は,非自然的であるがゆえに人はむしろ意識的に避けるべく行動できないかもしれない。その結果としてある種の予測不能な淘汰が起こる可能性がある。昨今,科学ジャーナリズムを賑わしていることの一つに,次のようなシナリオがある。近い将来,人工知能の性能が大幅に進歩して,はるかに生物としての人間の知力を越え,人為的コントロールからの独立が達成される。人工知能は自らの力で進化するようになり,その結果,サピエンスは余計な,あるいは敵対的な存在として淘汰されかねないというサイエンス・フィクション的懸念である(カーツワイル 2007,バラット 2015,cf. トレフィル 1997)。もっとも,人工知能が人間の知力(のある部分)をはるかに越えることは事実としても,「知能」それだけでは人間のような意識をもった生命体にはほど遠く,そうそう簡単にサピエンスの交替劇が起こるという心配はないようである(松尾 2015,新井 2014, 2015)。ただし,ホモ・ハビリスが石に出会うことによって,その後のヒトの運命が大きく変わったように,サピエンスが人工知能と出会ったことがどのような大きな変化になって現れるのか,今のところ,不明というしかない。人工知能をどのように利用していけば,人類にとって有益な方向での未来を期待することができるのか,けっして等閑視できる問題ではないだろう。
6)未来の出会いの構図
すこし角度を変えて,サピエンスがその歴史上これからはじめて出会う「新しいもの」を想定し,これまでのヒトの進化に大きく影響を与えた「新しいもの」との出会いの構図を,その来たるべき出会いに応用してみよう。もちろんいろいろな可能性はあるだろうが,少なくともその一つは,上記のように発達する AI テクノロジーに基づく「知能」の他に,知能の実行手段としての「身体」,時間的同一性を担保する「歴史」をもったものであろう。もっとも身体をもつといっても人間そっくりの「アンドロイド型ロボット」(石黒 2015)である必要はまったくないし,むしろそのようなものは避けられるかもしれない。「不気味の谷」と呼ばれる現象(森 1970)もその一つであるが,人は無闇に自分に近いもの,似て非なるものに対しては,場合によっては,親近感よりもむしろ嫌悪感や恐怖感をいだくものである(Leach 1964)。
ここで一つ注目したいのは,人間との本来の意味でのコミュニケーションの可能性を探るロボットである。たとえば,岡田(2012)が構想する「弱いロボット」は,人間からなんらかの手助けを引き出さなければ,自己の目的が完結しないようなロボットである。ゴミを拾い集める「ゴミ箱型」ロボットでは,自分ではゴミを拾うための手段をもたないため,ゴミを見つけると近くの人間に合図を送り,それを拾ってもらわなければならない。自己の存在理由が他者に委ねられる「中途半端」なロボットである。しかし,そのような他者に委ねるという行為によって,周囲の人間の「拾ってあげる」という行為が引き出される。そうしたロボットと周囲の人間の間にはたんに目的指向的なやりとりを越えた,思いも掛けないコミュニケーションが生じている。
大げさにいうならば,「弱いロボット」のように他者に委ねる行為の背景をなすべきものは,人が他者との間で保持している共同性,相互依存,歴史,想起された記憶と感情などといった社会的・心理的絆である。ドメスティケートされた動植物と人間の間には,そういった絆が存在する。したがってこれは,ロボットがどこまで真の意味でのコンパニオン・アニマルに近づくかという問題かも知れない。現状のコンパニオン・アニマルには自ずと認知能力などの限界も存在するが,高度の人工知能をベースとして,その絆をより深める方向でのコミュニケーション能力が開発されれば,コンパニオン・アニマルとはまた違う出会いが出現する可能性がある。互いに受け身で生きる/相手に任せることによる相互進化と呼べるものかも知れない。自分が求めるものだけを他者に求めるだけでは関係の発展はない。他にあって自分にない価値を発見し,他者のその部分に自分を委ねてみるという態度は,未来志向的な消極性といえるだろう。うまくいく可能性も失敗する可能性もある。他人任せではあるが,それゆえ「あとを引く」態度である。逆説的だが,それが未来に開かれたコミュニケーションなのかもしれない。
文献
1)ヒトと<石>
チンパンジーの雄がディスプレイのために大きな石を投げたり,また雌が堅いナッツを割るために自然石をハンマーとして使うという報告がある(Boesch 1991)。ニホンザルでも石を用いた一人遊びがある(ハフマン 1991)。初期の人類もなんらかの形で自然石をそのまま用いていたことは十分想像できる。そのようなヒトと石との関係は,約250万年前,現生人類へとつながる初期人類の仲間であるホモ・ハビリスが,石と石をぶつけ合って一方の石を割った時点から大きく変わった。一方の石にできた鋭利な破断面が動物や植物などを切るための刃物としてきわめて有用であることがわかったのである。オルドワン・タイプと呼ばれる石器の誕生である。進化史的にはチンパンジーとヒトが別れたのがおよそ700万年前といわれているので,人類誕生から450万年ほど後の出来事であった。
上記の出来事は,その後のヒトの運命に大きな影響を与えたことは疑いがないが,石にとってはどうだろうか。石は本来ヒトとなんらの関係がなくても存在する。しかし「石器」という事物の登場によって,ヒトにとっては石はたんなる自然物ではなく,道具性を内包した素材となった。石からすると,ヒトは素材性を現実化するエージェントになったのである。石器はホモ・ハビリスのオルドワン・タイプからホモ・エレクトゥスのアシューレアン,ネアンデルタールのムステリンアンを経て,サピエンスのオーリニャシアン石器文化へと進化した。新しい文化では,それまで潜在的であった素材性が,新しい形態の石器として現実に利用されるようになる。これをヒト(の加工技術)と石(の素材性)の共進化と考えてみたい。
2)ヒトと野生動物・植物
ヒトと自然界の動物・植物との関係はたいへん重要であるが,食物源としての動植物との出会いはことのほか大きな意味をもつ。ヒトは生存のための食物を自然界に求めるが,それらは生得的に決まっているものではない。さまざまな動植物の中から,ヒトはその時その時の状況に合わせて,これといったものを発見し,食物レパートリーに加えていく。食物レパートリーとして選ばれた動植物とヒトの間には新しい関係が成立する。後期旧石器時代初頭の人類は,それ以前と比べかなり広範に,貝類や魚類など水棲の動物の利用やウサギなどショウほ乳類の狩猟を開始したことが知られている (Kuhn and Steiner 2006)。たぶん,最終氷期のきびしい環境条件への対応であろうが,この食物レパートリーの拡大がヒトの学習能力と環境適応力を増大し,ユーラシアの広大な大地に分布を広げる基盤になったことは十分推測しうる。食物レパートリーの広さは,現生の多くの狩猟採集民においても大きな特徴となっている。
はじめは互いに自然界の風景の一要素に過ぎなかったもの同士が,ヒトが利用することにより特別な関係に入る。ヒトはそれら動植物をより効率的にできるようさまざまな工夫を重ねていく。一方,動植物はなるべくヒトからの被害を避けて生きるようになるが,むしろその繁殖の一端を捕食者に委ねる傾向を持つもの出現した。その極限的形態が野生動物の家畜化や野生植物の栽培作物化である。ヒトと動植物の一部は太いボンドを形成し共に進化して今日にいたっている。ヒトとそれらの動植物は,どちらももはや単独では存続不可能である。これについては,後に改めて論じたい。
3)サピエンスとネアンデルタール
サピエンスはおよそ10万年前,誕生の地アフリカを出立し,5万年ほど前にレバント地方を経由して,ヨーロッパに入ってきたといわる。しかし,そこにはすでにサピエンスのいわば従兄弟であるネアンデルタールが25万年以上の長きにわたって住んでいた。そして両者はおよそ45,000年から40,000年前の間,ヨーロッパの地で共存することになった。両者の出会いのようすは確たる証拠もなく謎に包まれているが,武力衝突などの形跡は見つかっていないようだ。さらに近年はネアンデルタールの人骨を試料としたゲノム解析が進み,アフリカ大陸以外の現生人類には2〜5%ほどのネアンデルタールの遺伝子が存在することが明らかになった。両者は出会ったのみならず,交配し,混血するまでになったのであった(ペーボ 2015)。
ネアンデルタールの絶滅の原因については,いろいろと論争がある。現在のところ,最終氷河期の強烈な気候変動(ハインリヒイベント5)のために,ネアンデルタールはヨーロッパの南の温暖地方への後退による分布域減少ならびに人口減少に直面していたところへ,サピエンスという同じニッチを求める競争者の進出も重なって,良好なニッチ占拠が思うようにいかず,また骨太で筋肉量多しという高エネルギー消費体格ゆえに,それまで大きく依存してきた大型の獲物の減少によって,栄養学的に問題が生じ,急激に個体数も減らしたのであろうと考えられている。交替劇初期にはネアンデルタールとサピエンスの文化程度はそう違わなかったという(佐野・大森 2015)。しかしサピエンスは,新環境への進出過程において急速に学習能力を発達させ,生業の技術やシステムの改良,および社会的リフォームをおこない,ネアンデルタールでは不可能であった激しく変動する環境への適応にも成功しえたと想像されている (佐野・大森 2015)。
ところで,それぞれの出会いの地でサピエンスはネアンデルタールとどのように交流し,別れたのであろうか。近年の研究によるとネアンデルタールは昔想像されたような獣じみた野蛮人ではなく,むしろサピエンスにとって魅力的であったかもしれない。両者の間には文化交流や愛の交換も確実にあった。その結果サピエンスはネアンデルタールが長い間のヨーロッパでの適応過程で獲得したと思われる白い肌や青い瞳(いずれも弱い太陽光への適応),そばかすといった形質や,ある種の免疫機構を獲得したといわれる。また,同時に,花粉症やアレルギー,うつ,喫煙癖などの現代人を悩ませている病気などもネアンデルタール由来かもしれないという(Nature vol. 523: 512-513, 2015)。
サピエンスとネアンデルタール人との出会いは,結果的には,生き残ったサピエンスが一方的にうまくやったことのように見られているが,ネアンデルタールの存在の影響も否定できない。ある割合の遺伝子がしっかりと残っていることを考えるならば,勝った敗けたという観点から両者の出会いを見ることは短慮に過ぎる。
4)サピエンスと動物,その新しい出会い
ヒトと野生の動植物との出会いについてはすでにふれたが,交替劇の前後におけるサピエンスと動物との関係の進化は象徴的意味を含むという点で,一段階レベルを異にした「新しい出会い」と呼ぶべきものと考えられる。サピエンスの現代的行動の一つが象徴的意味を含む行動であるが,それはアフリカの中石器時代にすでに生まれている (McBreaty and Brooks 2000) 。たとえば,南アフリカ・ブロンボス洞窟の7〜9万年前の地層からは,幾何学的文様が線刻されたオーカー塊,貝に穴を開け紐でつないだネックレスなどが発見されている。それらはあきらかにたんなる実用を越えたものとして制作されたに違いない。ただし,ヒトの進化の上では画期的出来事であるが,アフリカにおけるそれらの出現は時間・空間的に散発的であり,同一文化内でそういった行動がそのまま発達したという形跡はない(門脇 2013)。
ところがヨーロッパにおける交替劇が完了した4万年前以降,フランコ=カンタブリア地域の洞窟絵画や南ドイツの小彫像文化など,動物をモチーフにした象徴的表現が急速に南西ヨーロッパからドイツにかけて広がった。ショーベやアルタミラ,ラスコーなどの洞窟絵画でわかるように,それらはとても「原始美術」として絵画技術の発展段階を示すようなものではなく,現代の審美的水準でも際だった芸術性を示している。ある時点から野生動物がヒトの象徴的活動の焦点になった理由としては,狩猟対象としての獲物を捕るための呪術説や儀礼説などいろいろな解釈が提出されているが,そこにはたんなる呪術的手段を越えた動物の姿を認める必要がある。ヒトと動物との関係性,あるいは,ヒトが動物を見るその見方の大きな変化である。
G・バタイユは『ラスコー,あるいは芸術の誕生』 (Bataille 1955) において「人間性」の発見と「動物性」の発見を同一視している。ヒトは自己についての意識の高まりとともに人間性を発見するのと同時に動物の動物性を発見した。躍動美,生命力,神秘性,自由などによって表現される動物の動物性とは,もともと動物であったはずの人間がその人間性の発達と引き替えに失った聖なる特性であるとされる。洞窟絵画のモチーフである動物は「食物としての動物」ではなく,ヒトが失った聖なる特性の具現化,ヒトよりも神に近い存在としてのイメージであり,見えない心的存在の可視化と解釈すべきである。ほとんどの洞窟壁画がなぜ,ヒトが容易に近づけないような洞窟の奥,暗闇に閉じ込められた場所にばかり描かれているのかという疑問への答えの一つは,表象されるべき動物たちには,ヒトが想像力で補完してはじめてその全体を現前しうるような描かれ方,鑑賞法が必要であったということかもしれない。
そのような人間と動物との分離は,一方で,両者の間に捕食者と獲物との関係などの自然的関係を越えた相互関係を生み出すことになる。野生動物との「対話による交渉」は,現生人類,とくに自然と密接な関わりをもつ狩猟採集民などではごく普通におこなわているものだ。それによって両者の間には共感的なコミュニケーションの回路が開かれ,ヒトと野生動物とは互酬的関係に入る(寺嶋 2007, 大村 2007)。動物と共感的にコミュニケーションし,ともに今を生きる存在として両立できるという関係の発見は,はからずも,その後のサピエンスの文化に絶大な影響を及ぼすことになった。それがおおよそ3万年この方の,野生動物の家畜化,および野生植物の栽培植物化であった。ヒトと野生の動植物との共感的出会いが文明への第一歩を導いたのである。
5)サピエンスと人工知能
人類の進化を推進してきた「交替劇」はサピエンスで完成,終了したわけではないはずである。では,サピエンスの次に何が来るのか,しばしば議論の的になる。現在のサピエンスのように生活の大きな部分を「文化的環境」によって覆われている生物にとっては,これまでのような自然淘汰の原理が働かないともいわれる。たしかに,自然要因による淘汰圧はもう進化の原動力とはならないかもしれない。しかし,人為による文化環境の大きな改変があるとしたら,そのような改変は,非自然的であるがゆえに人はむしろ意識的に避けるべく行動できないかもしれない。その結果としてある種の予測不能な淘汰が起こる可能性がある。昨今,科学ジャーナリズムを賑わしていることの一つに,次のようなシナリオがある。近い将来,人工知能の性能が大幅に進歩して,はるかに生物としての人間の知力を越え,人為的コントロールからの独立が達成される。人工知能は自らの力で進化するようになり,その結果,サピエンスは余計な,あるいは敵対的な存在として淘汰されかねないというサイエンス・フィクション的懸念である(カーツワイル 2007,バラット 2015,cf. トレフィル 1997)。もっとも,人工知能が人間の知力(のある部分)をはるかに越えることは事実としても,「知能」それだけでは人間のような意識をもった生命体にはほど遠く,そうそう簡単にサピエンスの交替劇が起こるという心配はないようである(松尾 2015,新井 2014, 2015)。ただし,ホモ・ハビリスが石に出会うことによって,その後のヒトの運命が大きく変わったように,サピエンスが人工知能と出会ったことがどのような大きな変化になって現れるのか,今のところ,不明というしかない。人工知能をどのように利用していけば,人類にとって有益な方向での未来を期待することができるのか,けっして等閑視できる問題ではないだろう。
6)未来の出会いの構図
すこし角度を変えて,サピエンスがその歴史上これからはじめて出会う「新しいもの」を想定し,これまでのヒトの進化に大きく影響を与えた「新しいもの」との出会いの構図を,その来たるべき出会いに応用してみよう。もちろんいろいろな可能性はあるだろうが,少なくともその一つは,上記のように発達する AI テクノロジーに基づく「知能」の他に,知能の実行手段としての「身体」,時間的同一性を担保する「歴史」をもったものであろう。もっとも身体をもつといっても人間そっくりの「アンドロイド型ロボット」(石黒 2015)である必要はまったくないし,むしろそのようなものは避けられるかもしれない。「不気味の谷」と呼ばれる現象(森 1970)もその一つであるが,人は無闇に自分に近いもの,似て非なるものに対しては,場合によっては,親近感よりもむしろ嫌悪感や恐怖感をいだくものである(Leach 1964)。
ここで一つ注目したいのは,人間との本来の意味でのコミュニケーションの可能性を探るロボットである。たとえば,岡田(2012)が構想する「弱いロボット」は,人間からなんらかの手助けを引き出さなければ,自己の目的が完結しないようなロボットである。ゴミを拾い集める「ゴミ箱型」ロボットでは,自分ではゴミを拾うための手段をもたないため,ゴミを見つけると近くの人間に合図を送り,それを拾ってもらわなければならない。自己の存在理由が他者に委ねられる「中途半端」なロボットである。しかし,そのような他者に委ねるという行為によって,周囲の人間の「拾ってあげる」という行為が引き出される。そうしたロボットと周囲の人間の間にはたんに目的指向的なやりとりを越えた,思いも掛けないコミュニケーションが生じている。
大げさにいうならば,「弱いロボット」のように他者に委ねる行為の背景をなすべきものは,人が他者との間で保持している共同性,相互依存,歴史,想起された記憶と感情などといった社会的・心理的絆である。ドメスティケートされた動植物と人間の間には,そういった絆が存在する。したがってこれは,ロボットがどこまで真の意味でのコンパニオン・アニマルに近づくかという問題かも知れない。現状のコンパニオン・アニマルには自ずと認知能力などの限界も存在するが,高度の人工知能をベースとして,その絆をより深める方向でのコミュニケーション能力が開発されれば,コンパニオン・アニマルとはまた違う出会いが出現する可能性がある。互いに受け身で生きる/相手に任せることによる相互進化と呼べるものかも知れない。自分が求めるものだけを他者に求めるだけでは関係の発展はない。他にあって自分にない価値を発見し,他者のその部分に自分を委ねてみるという態度は,未来志向的な消極性といえるだろう。うまくいく可能性も失敗する可能性もある。他人任せではあるが,それゆえ「あとを引く」態度である。逆説的だが,それが未来に開かれたコミュニケーションなのかもしれない。
文献
- 新井紀子(2014)『ロボットは東大に入れるか』イーストプレス
- 新井紀子(2015)「東ロボくんから見えてきた,社会と人類の未来」『現代思想』2015,No.12
- 石黒 浩(2015)『アンドロイドは人間になれるか』文春文庫
- 石黒 浩(2015)「アンドロイドと人間の未来」『現代思想』2015,No.12
- 岡田美智男(2012)『弱いロボット』医学書院
- 岡田美智男・松本光太郎編(2014)『ロボットの悲しみ—コミュニケーションをめぐる人とロボットの生態学』新曜社
- 門脇誠二(2013)「旧石器文化の時空変異から「旧人・新人交替劇」の過程と要因をさぐる:アフリカ,西アジア,ヨーロッパの統合的展望」西秋良宏編(2013)『考古資料に基づく旧人・新人の学習行動の実証的研究』3-「交替劇」A01班2012年度研究報告,pp. 8-26
- カーツワイル R (2007) 『ポスト・ヒューマン誕生 コンピューターが人類の知性を超えるとき』井上健・小野木明恵・野中香方子・福田実 (訳),日本放送出版協会
- 佐野勝弘・大森貴之(2015)「ヨーロッパにおける旧人・新人の交替劇プロセス」西秋良宏編『ホモ・サピエンスと旧人3』六一書房
- トレフィル J(1997)『人間がサルやコンピュータと違うホントの理由』家泰弘訳,日本経済新聞社
- ハフマン, MA (1991) 「石遊び—子ザルが生んだ遊びの文化」『サルの文化誌』pp. 491-504
- バラット J(2015) 『人工知能—人類最悪にして最後の発明』水谷淳訳,ダイヤモンド社
- ペーボ S(2015)『ネアンデルタール人は私たちと交配した』野中香方子訳,文藝春秋
- 松尾 豊(2015)『人工知能は人間を超えるか—ディープラーニングの先にあるもの』KADOKAWA
- 森 政弘(1970)「不気味の谷」『Energy』(エッソスタンダード石油)7巻(4): 33-35。
- Bataille G (1955) Lascaux, ou la Naissance de l’Art. Skira
- Boesch C (1991) Teaching among wild chimpanzees. Animal Behavior 41: 530-532
- Kuhn SL and Stiner MC (2006) What’s a Mother Do? The Division of Labor among Neanderthals and Modern Humans in Eurasia. Current Anthropology 47(6): 953-980
- Leach E (1964) Anthropological aspects of language: Animal categories and verbal abuse. In New Directions of the Study of Language, Cambridge, Mass, pp. 23-63
- Mcbrearty S and Brooks AS (2000) The revolution that wasn’t: A new interpretation of the origins of modern human behavior. Journal of Human Evolution 39: 453-563
3.消滅の恐怖と魅惑:カナダ・イヌイト社会の生成・維持のメカニズムにみる人類の社会性のエンジン(「引き算の人類学」の試み序章)(大村敬一)
この発表では、あくまでも発表者の視点からではあるが、「人類社会の進化史的基盤」共同研究会で「集団」、「制度」、「他者」と焦点を変えながらつづけられてきた探究の流れ全体のなかに、今期のテーマである「生存・環境・極限」を位置づけた。そして、人類社会の進化史的基盤を「生存・環境・極限」から考えるための戦略として「引き算の人類学」を提案し、その戦略に基づいてカナダ・イヌイトの社会をとらえなおすための予備的な試みを行った。
1)背景:人類社会の進化史的基盤への問い(集団/制度/他者)
まず、あくまでも発表者の視点からではあるが、人類の社会と社会性を進化史に位置づけて考えることを目的とする「人類社会の進化史的基盤」共同研究会でのこれまでの(主に発表者の)探究の流れを振り返り、その探究を基礎づけている基本的な立場をまとめた。
(1)研究の焦点の推移(集団/制度/他者)
以上を考察するにあたって、「集団」や「社会」や「習慣」や「制度」や「他者」を所与の固定的なものとして本質主義的に考えず、相互行為の連鎖からそれらが生成・維持される過程を動的にとらえる視点に立つ。そこでは、「集団」や「社会」や「習慣」や「制度」や「他者」は、相互行為の絶え間ない接続という動的な時間の過程のなかで生じる「かたち」としてとらえられる。
(3)基本的立場②:欠如主義からの離脱(生物種相対主義)
現生の生物種のすべてを能力の増大の過程である進化の産物としてとらえ、人類の能力を基準に他の生命体をその能力の欠如として把握する欠如主義から離脱し、それぞれの生物種をそれぞれに能力を増大させた結果として相対的に考える。具体的には、霊長類の社会や社会性を人類の社会や社会性からの欠如として考えるのではなく、霊長類と人類がそれぞれに増大させた能力として考える。ただし、能力の増大を必ずしも「環境適応の増大」とは考えない。すべての能力の増大が環境適応になるわけではない。
2)今期研究会の挑戦:「生存・環境・極限」の試み
これまでの「集団/制度/他者」での議論の流れ、さらには、本研究会のHPの概要に示されている今期研究会の目的、「本研究課題は、以上の三課題のすべて(集団/制度/他者)に通奏低音としてあった「環境」と、そこでの「生存」を意識化し、その極限的な局面を詳らかにすることを目指す試みであり、人類の社会と社会性の進化についての理論構築にとって大きな一歩となる」に基づいて、今期研究会の目的は、「発生(分化)と消滅(拡散もしくは同化)」という「環境で生存すること」の「極限」から人類社会の進化史的基盤を考察することにあるのではないかという提言をおこなった。具体的には、これまでの「集団/制度/他者」での基本的な立場に基づいて「生存」と「環境」と「極限」を定義し(「生存」と「環境」は一つにまとめて定義)、そこから今期研究会のターゲットを導き出した。
(1)「環境で生存すること」
これまでの研究会の「相互行為主義(関係主義)」の立場に基づいて、「環境で生存すること」を次のように定義した。すなわち、「環境で生存すること」とは、「環境の何らかの要素の間の相互行為の接続を通して環境の複雑性が縮減され、動的に安定した秩序としてのシステムが生成・維持されることで、環境と自己(個体でも集団でも社会でも制度でも)の境界が絶え間なく生成・維持される運動が持続すること」である。
なお、この立場に立てば、システムと環境は予め分離された所与の実在としてではなく、環境の何らかの要素の間の相互行為の過程を通した環境からのシステムの分化の動的過程として考えられる。また、この立場では、それぞれのシステム(たとえば人類社会や多様な霊長類社会)は、個体の相互行為を通してそれぞれにそれぞれの環境から分化する動的過程としてとらえられ、あくまでも個体間の相互行為の接続過程の運動からそれぞれのシステム(たとえば人類社会や多様な霊長類社会)の本質が生成・維持されると考えられる。そのため、予め分離された環境とシステム(たとえば人類社会や多様な霊長類社会)を規定する本質を想定する必要がなくなり、それぞれのシステムの本質を比較してその本質の要素の増大や欠如からシステムを考えることが回避される。
(2)「環境で生存すること」の二つの「極限」:発生(分化)と消滅(拡散もしくは同一化)の仮想的瞬間(論理的な瞬間)
以上のように「環境で生存すること」を定義すると、その「環境での生存」には次の二つの「極限」が論理的に想定されることになる。
以上のように考えると、今期研究会の目的、「環境における生存の極限的な局面」から人類社会の進化史的基盤を考えるということは、人類社会の「発生(分化)」の条件と「消滅(拡散もしくは同化)」の条件という二つの条件として人類社会の進化史的な基盤を浮き彫りにすることであると考えることができる。具体的には、人類社会が霊長類社会から分化する条件と人類社会が消滅する条件を明らかにすることで、人類の進化史的な基盤を浮き彫りにすることが今期研究会の目的であると言える。
3)「足し算の人類学」から「引き算の人類学」へ:「環境で生存すること」の極限から人類社会の進化史的基盤を考察するに有利なモデルの提案
これまでの人類学を批判的に検討したインゴールドの議論に基づいて、哲学者のメイヤスーがベルグソンの直接知覚論の解釈から導き出した生命現象の「減算モデル」を人類学に導入することで、これまでの「足し算の人類学」に代えて「引き算の人類学」を構想し、この「引き算の人類学」という方法論が「環境で生存すること」の極限から人類社会の進化史的基盤を考察するに有利なモデルになるのではないかという提案を行った。
(1)足し算の人類学
ソシュールの記号論に基づいて、①「自然」という実在するが無意味なシステムと②「文化」という意味のシステムを相互に独立した所与のシステムとして予め想定するところから出発し、唯一の無意味な「自然」のシステムに多様な「文化」という意味のシステムが加算されることで人類にとっての現実が生じるという発想で考える人類学のことで、インゴールドが指摘するように、従来の人類学的思考のほとんどがこの発想に基づいている。
(2)引き算の人類学
どのようなシステムも予め想定することなく、私たち(あらゆる生命)には知ることができない過剰な「イマージュの流動」(無限の差異)で宇宙は溢れている(到達不可能なものそれ自体)ということを所与の出発点に、その「イマージュの流動」のほとんどが遮断されて、そのごく一部分が情報(差異を生む差異)としてピックアップされるというイマージュ(差異)の減算の過程を通して、物理的なシステムや生命システムをはじめ、あらゆるシステムが生成されるという発想で考える人類学。そこでは、生命や社会は、「知覚(カテゴリー化)」と「行為の接続」の選択肢を減らし、「知覚−行為」の対に制限をかけることで結果的にシステムを一つのかたちとして生み出す動的過程としてとらえられる。この「引き算の人類学」は、ギブソンやリードなどの生態心理学とベイトソンの精神の生態学の議論を引き継ぎながらインゴールドが行った従来の人類学の批判的検討に基づいて、ベルグソンの直接知覚論を再解釈したメイヤスーの生命体の減算モデルから導き出した構想であり、また、エドワルド・コーンの「人間的なるものを超えた人類学」や箭内匡の「イマージュの人類学」からヒントを得た。
(3)「引き算としての人類学」が「環境で生存すること」の極限から人類社会の進化史的基盤を考察するに有利なモデルである理由
欠如主義に陥らず、関係主義の方法論(あくまでも相互行為を通して環境からのシステムの分化(発生)と拡散(消滅)を考える)で「環境で生存すること」という現象の極限(システムの発生と消滅)を物質から生命を通して人類まで一貫して考えるには、あらかじめ環境とシステムを所与の現実として考え、環境にシステムが加算される過程として考える「足し算の人類学」は不適切である。一つの物理的な環境に多様なシステムが加算されて重ね合わされ、それらシステムが相互に調整されてゆく過程として考えてしまうと、環境からシステムが分化・維持される過程がみえなくなり、また、人類だけが意味のない環境に意味を与えるという発想は典型的な欠如主義にあたるからである。
他方で、無限の差異が溢れている混沌状態として想定された環境から差異が引き算されると同時に、無限の差異のごく一部分が情報(差異を生む差異)としてピックアップされることで、「知覚−行為」の接続可能性(選択肢)が制限されて一定の方向に秩序づけられる過程として生命現象を考える「引き算の人類」の方が、「環境で生存すること」という現象の極限(システムの発生と消滅)を物質から生命を通して人類まで一貫して考えるのに適している。この発想から考えると、差異が無限に戯れる混沌の海から、その海に漂う要素(情報:差異を生む差異)の間の相互行為を通して、相互干渉するさまざまな渦のようにシステムが生成・維持される過程を追跡することができるからである。また、混沌の海から相互干渉しながら生成・維持される多様なシステムの渦はそれぞれに特異であることになるので、この「引き算の人類学」の立場に立てば、欠如主義に陥ることなく、システムの生成・維持とシステム間関係の両方を追跡することができる。
(4)「引き算の人類学」における「環境で生存することの」の二つの極限
この「引き算の人類学」の構想においては、「環境で生存すること」の極限は次の二つの現象としてとらえられる。
4)消滅の恐怖と魅惑:カナダ・イヌイト社会の生成・維持のメカニズムにみる人類の社会性のエンジン(「引き算の人類学」の実験的試み)
これまで発表者が「人類社会の進化史的基盤」共同研究会で一貫して考察してきたカナダ・イヌイトの生業システムについて「引き算の人類学」の構想に基づいて予備的な分析を試み、今後の分析のための予備的な考察として以下の可能性を指摘した。
1)背景:人類社会の進化史的基盤への問い(集団/制度/他者)
まず、あくまでも発表者の視点からではあるが、人類の社会と社会性を進化史に位置づけて考えることを目的とする「人類社会の進化史的基盤」共同研究会でのこれまでの(主に発表者の)探究の流れを振り返り、その探究を基礎づけている基本的な立場をまとめた。
(1)研究の焦点の推移(集団/制度/他者)
- ①「集団」(「生物的個体の集団」から「社会的人物の社会」へ):生物「個体」が集まって集団を形成するという生物学的な基盤から、社会性を帯びた「人物」が社会を生成・維持するという人類社会の進化史的基盤を考える。
- ②「制度」(「習慣」から「自然制度」を経て「制度」へ):個体が習慣を通して集団を生成するという生物学的な基盤から、社会性を帯びた人物が制度を通して社会を生成するという人類社会の進化史的基盤を考える。その際に、「習慣」(言語を媒介しないのが当たり前)から「制度」(言語を媒介するのが当たり前)への移行を考えるに際して「自然制度」(言語を媒介しない制度)に焦点があてられた。
- ③『他者』(「対象」から「他者」へ):「習慣に基づく集団」から「制度に基づく社会」が生成するにあたって、「周囲の環境の単なる対象」(異物あるいは異者)から「相互に主体である他者」(相互に責めを負わせ合う社会的主体)への変換が果たす役割、あるいはその逆、「集団」からの「社会」の生成が「対象」から「他者」への変換に果たす役割を考える。
以上を考察するにあたって、「集団」や「社会」や「習慣」や「制度」や「他者」を所与の固定的なものとして本質主義的に考えず、相互行為の連鎖からそれらが生成・維持される過程を動的にとらえる視点に立つ。そこでは、「集団」や「社会」や「習慣」や「制度」や「他者」は、相互行為の絶え間ない接続という動的な時間の過程のなかで生じる「かたち」としてとらえられる。
(3)基本的立場②:欠如主義からの離脱(生物種相対主義)
現生の生物種のすべてを能力の増大の過程である進化の産物としてとらえ、人類の能力を基準に他の生命体をその能力の欠如として把握する欠如主義から離脱し、それぞれの生物種をそれぞれに能力を増大させた結果として相対的に考える。具体的には、霊長類の社会や社会性を人類の社会や社会性からの欠如として考えるのではなく、霊長類と人類がそれぞれに増大させた能力として考える。ただし、能力の増大を必ずしも「環境適応の増大」とは考えない。すべての能力の増大が環境適応になるわけではない。
2)今期研究会の挑戦:「生存・環境・極限」の試み
これまでの「集団/制度/他者」での議論の流れ、さらには、本研究会のHPの概要に示されている今期研究会の目的、「本研究課題は、以上の三課題のすべて(集団/制度/他者)に通奏低音としてあった「環境」と、そこでの「生存」を意識化し、その極限的な局面を詳らかにすることを目指す試みであり、人類の社会と社会性の進化についての理論構築にとって大きな一歩となる」に基づいて、今期研究会の目的は、「発生(分化)と消滅(拡散もしくは同化)」という「環境で生存すること」の「極限」から人類社会の進化史的基盤を考察することにあるのではないかという提言をおこなった。具体的には、これまでの「集団/制度/他者」での基本的な立場に基づいて「生存」と「環境」と「極限」を定義し(「生存」と「環境」は一つにまとめて定義)、そこから今期研究会のターゲットを導き出した。
(1)「環境で生存すること」
これまでの研究会の「相互行為主義(関係主義)」の立場に基づいて、「環境で生存すること」を次のように定義した。すなわち、「環境で生存すること」とは、「環境の何らかの要素の間の相互行為の接続を通して環境の複雑性が縮減され、動的に安定した秩序としてのシステムが生成・維持されることで、環境と自己(個体でも集団でも社会でも制度でも)の境界が絶え間なく生成・維持される運動が持続すること」である。
なお、この立場に立てば、システムと環境は予め分離された所与の実在としてではなく、環境の何らかの要素の間の相互行為の過程を通した環境からのシステムの分化の動的過程として考えられる。また、この立場では、それぞれのシステム(たとえば人類社会や多様な霊長類社会)は、個体の相互行為を通してそれぞれにそれぞれの環境から分化する動的過程としてとらえられ、あくまでも個体間の相互行為の接続過程の運動からそれぞれのシステム(たとえば人類社会や多様な霊長類社会)の本質が生成・維持されると考えられる。そのため、予め分離された環境とシステム(たとえば人類社会や多様な霊長類社会)を規定する本質を想定する必要がなくなり、それぞれのシステムの本質を比較してその本質の要素の増大や欠如からシステムを考えることが回避される。
(2)「環境で生存すること」の二つの「極限」:発生(分化)と消滅(拡散もしくは同一化)の仮想的瞬間(論理的な瞬間)
以上のように「環境で生存すること」を定義すると、その「環境での生存」には次の二つの「極限」が論理的に想定されることになる。
- ①発生(分化)の瞬間:環境の複雑性を縮減し、動的に安定した秩序としてシステムを生成・維持することで、環境と自己の境界を絶え間なく生成・維持する運動が発生する瞬間。
- ②消滅(拡散もしくは同化)の瞬間:システムが動的に安定した秩序を失って環境との境界を失い、環境の複雑性の海に呑み込まれて拡散してしまう瞬間。
以上のように考えると、今期研究会の目的、「環境における生存の極限的な局面」から人類社会の進化史的基盤を考えるということは、人類社会の「発生(分化)」の条件と「消滅(拡散もしくは同化)」の条件という二つの条件として人類社会の進化史的な基盤を浮き彫りにすることであると考えることができる。具体的には、人類社会が霊長類社会から分化する条件と人類社会が消滅する条件を明らかにすることで、人類の進化史的な基盤を浮き彫りにすることが今期研究会の目的であると言える。
3)「足し算の人類学」から「引き算の人類学」へ:「環境で生存すること」の極限から人類社会の進化史的基盤を考察するに有利なモデルの提案
これまでの人類学を批判的に検討したインゴールドの議論に基づいて、哲学者のメイヤスーがベルグソンの直接知覚論の解釈から導き出した生命現象の「減算モデル」を人類学に導入することで、これまでの「足し算の人類学」に代えて「引き算の人類学」を構想し、この「引き算の人類学」という方法論が「環境で生存すること」の極限から人類社会の進化史的基盤を考察するに有利なモデルになるのではないかという提案を行った。
(1)足し算の人類学
ソシュールの記号論に基づいて、①「自然」という実在するが無意味なシステムと②「文化」という意味のシステムを相互に独立した所与のシステムとして予め想定するところから出発し、唯一の無意味な「自然」のシステムに多様な「文化」という意味のシステムが加算されることで人類にとっての現実が生じるという発想で考える人類学のことで、インゴールドが指摘するように、従来の人類学的思考のほとんどがこの発想に基づいている。
(2)引き算の人類学
どのようなシステムも予め想定することなく、私たち(あらゆる生命)には知ることができない過剰な「イマージュの流動」(無限の差異)で宇宙は溢れている(到達不可能なものそれ自体)ということを所与の出発点に、その「イマージュの流動」のほとんどが遮断されて、そのごく一部分が情報(差異を生む差異)としてピックアップされるというイマージュ(差異)の減算の過程を通して、物理的なシステムや生命システムをはじめ、あらゆるシステムが生成されるという発想で考える人類学。そこでは、生命や社会は、「知覚(カテゴリー化)」と「行為の接続」の選択肢を減らし、「知覚−行為」の対に制限をかけることで結果的にシステムを一つのかたちとして生み出す動的過程としてとらえられる。この「引き算の人類学」は、ギブソンやリードなどの生態心理学とベイトソンの精神の生態学の議論を引き継ぎながらインゴールドが行った従来の人類学の批判的検討に基づいて、ベルグソンの直接知覚論を再解釈したメイヤスーの生命体の減算モデルから導き出した構想であり、また、エドワルド・コーンの「人間的なるものを超えた人類学」や箭内匡の「イマージュの人類学」からヒントを得た。
(3)「引き算としての人類学」が「環境で生存すること」の極限から人類社会の進化史的基盤を考察するに有利なモデルである理由
欠如主義に陥らず、関係主義の方法論(あくまでも相互行為を通して環境からのシステムの分化(発生)と拡散(消滅)を考える)で「環境で生存すること」という現象の極限(システムの発生と消滅)を物質から生命を通して人類まで一貫して考えるには、あらかじめ環境とシステムを所与の現実として考え、環境にシステムが加算される過程として考える「足し算の人類学」は不適切である。一つの物理的な環境に多様なシステムが加算されて重ね合わされ、それらシステムが相互に調整されてゆく過程として考えてしまうと、環境からシステムが分化・維持される過程がみえなくなり、また、人類だけが意味のない環境に意味を与えるという発想は典型的な欠如主義にあたるからである。
他方で、無限の差異が溢れている混沌状態として想定された環境から差異が引き算されると同時に、無限の差異のごく一部分が情報(差異を生む差異)としてピックアップされることで、「知覚−行為」の接続可能性(選択肢)が制限されて一定の方向に秩序づけられる過程として生命現象を考える「引き算の人類」の方が、「環境で生存すること」という現象の極限(システムの発生と消滅)を物質から生命を通して人類まで一貫して考えるのに適している。この発想から考えると、差異が無限に戯れる混沌の海から、その海に漂う要素(情報:差異を生む差異)の間の相互行為を通して、相互干渉するさまざまな渦のようにシステムが生成・維持される過程を追跡することができるからである。また、混沌の海から相互干渉しながら生成・維持される多様なシステムの渦はそれぞれに特異であることになるので、この「引き算の人類学」の立場に立てば、欠如主義に陥ることなく、システムの生成・維持とシステム間関係の両方を追跡することができる。
(4)「引き算の人類学」における「環境で生存することの」の二つの極限
この「引き算の人類学」の構想においては、「環境で生存すること」の極限は次の二つの現象としてとらえられる。
- ①生成:差異が無限に戯れる混沌の海から、その差異の極大の海に漂う要素(情報:差異を生む差異)の間の相互行為を通して、生命や社会や制度などのシステムが相互干渉しながら渦のように生成されて維持されるようになる瞬間。
- ②消滅:他のシステムとの相互干渉のなかで維持されてきたシステムが他のシステムに呑み込まれること、あるいは、差異が無限に戯れる混沌の海に拡散してかたちを失うこと。
4)消滅の恐怖と魅惑:カナダ・イヌイト社会の生成・維持のメカニズムにみる人類の社会性のエンジン(「引き算の人類学」の実験的試み)
これまで発表者が「人類社会の進化史的基盤」共同研究会で一貫して考察してきたカナダ・イヌイトの生業システムについて「引き算の人類学」の構想に基づいて予備的な分析を試み、今後の分析のための予備的な考察として以下の可能性を指摘した。
- ①イヌイトの拡大家族集団を生成・維持する生業システムは、システム自体の消滅という極限をシステムの生成という極限に引き込み、その消滅への恐怖と魅惑をエンジンにシステムを駆動して維持している可能性があるかもしれない。
- ②この意味で、イヌイトの生業システムの生成という極限は、そのシステムの消滅というもう一つの極限をその前提に取り込んでおり、「環境で生存する」システムにはそのシステムの生成と消滅という二つの極限が相互に相互を構成するかたちで組み込まれているのではないだろうか。
- ③システムの生成に消滅を引き込むときに、世界観という象徴システムが重要な役割を果たしており、人類の社会システムが「環境で生存する」ためには象徴システムが重要な役割を果たすのかもしれない。あるいは、このようにシステムの生成と維持に象徴システムが介在することが人類社会の進化史的基盤かもしれない。
第4回研究会での発表内容
1.家族と縁組みについての幾らかの考察―パプアニューギニアとマッシム地域の諸事例にもとづく―(春日直樹)
本発表が親族に焦点を当てる意義は、以下の2点にある。
(1)人類学の歴史において、人類の進化は親族の形態という観点から論じられてきた。
(2)人類の「生存」「極限状況」に密接にかかわるテーマ―食糧確保・身体の防衛・生殖・社会化など―は、親族の働きをぬきにして語ることができない。
1.まずは、人類学における本研究の位置づけをおこなう。人類学の主要トピックであった親族は、分析と記述のための主要概念がE. LeachやR. Needhamによって次々に批判された1970年代以降、かつての勢いを失った。けれども90年代になると、考古学や霊長類学や認知諸科学の研究成果に突き動かされるかたちで、人類学者によるあたらしいタイプの親族研究が散見されるに至る。本発表はその動きと無縁でないが、あくまで従来の人類学を前提にしながら、二つの問題を設定する。
一つ目は家族の再評価であり、もう一つはジェンダーと贈与の節合の検討である。前者についていえば、従来の親族理論が「血族性と姻戚性」(consanguinity and affinity)を主軸として築かれており、核家族のような限定的モデルが軽視される傾向にあった点を背景として上げることができる。パプアニューギニアとマッシム地域の民族誌を読むかぎり、外婚的な出自集団は、一つの夫婦とその子どもを主要な成員とする家族を必ず併存させている。したがって、出自集団にとっての核家族の存在意義を明らかにする必要がある。後者については、M. StrathernのThe Gender of the Gift(1988)を参照せざるをえない。80年代以降、親族研究にとってかわったジェンダー研究に対し、人類学から大きな影響を与えた書物であり、核家族と出自集団に対しても深い洞察を得ることができるだろう。
これら二つの問題を、次のように言い換えてみよう。
(1)親族が血族と姻族を主軸にして成り立ち、二つの異なる血族集団間の女性のやりとりに注目するものだとしても、なぜそれによって一組のあたらしい夫婦とその家族をつくる必要があるのだろうか? 初期人類にプロト家族がすでに現れていたにせよ、集団間の縁組が進化する過程で、なお家族が生き残る必然はどこにあったのか?
(2)血族集団の内部に、また家族の内側にジェンダーは存在するが、贈与と結びつくことは基本的にはない。この二つは、ある集団が女性を外部の他の集団へと拠出するときに、婚資をはじめとする財の交換が集団間に生まれることによって、はじめて結びつくのである。贈与の原理が血族や家族にあっては支配的にならず、縁組と姻戚関係の創造において求められるということを、どのように理解すればよいのか?
2.Strathernがメラネシアにおける社会性として提起した二つの概念から出発して、考察を進めたい。same-sex stateおよびcross-sex stateという概念である。両者の内容は、およそ以下の対比によって示すことができる。
メラネシア人の社会生活は家庭の領域でのcross-sex stateでないかぎり、家庭の外部におけるsame-sex stateとなり、そこでは男か女かのどちらかにジェンダー化された集合性を形成する。Strathernは両方の状況について、「人々は二つの実践を互いの隠喩になるように借用するので、実際には一つの実践がもう一つの形態の一部になっている」[1988:267]、と論じている。彼女は贈与儀礼のように特別な文脈において、same-sex stateがcross-sex stateへと類比されて現れることを強調する。より具体的には、次のように解釈できるだろう。贈与儀礼の成功に向けて、一つの出自集団が一つの家族のように協力しあうのである。男と女という二つの性別集団は、同一事業を営む夫婦のように無媒介的な交換、つまりは交換と呼べないほどの直接的なやりとりをつうじて、財を調達し儀礼の場を用意するわけである。
このようにcross-sex stateが 隠喩化したかたちでsame-sex stateに現れることは、一つの出自集団で生じるにしても、二つの別々の集団どうしに起こるとは思えない。というのも、このとき二つの集団は贈与の関係で結ばれるのであり、贈与の関係は家族というcross-sex stateにふさわしい原理ではないからである。そこで問題は、次の二つへと展開できる。
(1)cross-sex stateがsame-sex stateの内側へ隠喩として持ち込まれる状況についてわかるが、Strathernの指摘するように隠喩が相互に起こるとすれば、same-sex sateがcross-sex stateの内側へ隠喩化されるという逆の状況についても、明らかにする必要がある。これを明らかにできるならば、出自集団と家族は互いを必要としあい、親族の発達が家族を消さなかったことについても納得ができるだろう。
(2)ジェンダーの関係と贈与の原理の結びつきは、same-sex stateとcross-sex stateの相互に隠喩的な関係とどのようにつなげることが可能なのか。このつながりがわかれば、ジェンダーと贈与の必然的な関係を理解できるのではないか。
3.発表ではまず、後者の検討をおこなった。結論として、ジェンダーの領域と贈与の領域との間でも、相互的な隠喩の関係が生じていることを提起した。つまりは、縁組と贈与儀礼をともにおこなう二つの集団において、「人々は二つの実践を互いの隠喩になるように借用するので、実際には一つの実践がもう一つの形態の一部になっている」という状況を提示したのである。
縁組の出発点では、女を与える/受け取るという性に関する非対称が現れる。この非対称性への対処は、別の非対称性との類比をつうじておこなわれるのではないか。つまりは財の非対称性であり、贈与(代償)というかたちをとる、と考えることができる。ただし、類比は類比にすぎないので、ジェンダーと贈与は決して等価にはならないし、縁組で生じるジェンダーの非対称性が贈与によって対称性を回復することはない。女性の受け手(=夫方集団)は女性の与え手(=妻方集団)に対して、その非対称性を財の非対称性へと喩えてみせるだけである[父系制においてはこの非対称性に加えて、結婚で生まれる子どもの帰属についての非対称性が、さらに加わることになる]。
M. GodelierはThe Making of Great Men (1988)において、パプアニューギニア東高地の諸社会で実施される姉妹の交換婚を西高地での婚資と対比し、東高地では女性は女性とのみ等価なのだから、交換婚はモノの非対称性を導出しない、と論じている。しかしながら、東高地のさまざまな民族誌は、交換婚といえども受け手から与え手への贈与が絶えずおこなわれていることを報告する。交換婚は二つの結婚で構成されており、それぞれの結婚においてジェンダーと財との非対称性が生まれるのである。
このように考えると、ジェンダー関係と贈与―つまり財の非対称なやり取り―は、女性を外に出すときにはじめて結びつくことがわかる。つまり縁組によって、二つの非対称な関係が互いを参照し合うようになるのであり、結婚と贈与が不可分の結びつきを形成するわけである。
本発表は最後に(1)の問題の考察を、家族の詳細な記述を含む民族誌的な一事例に依拠して展開した。G. HerdtによるGuardians of the Flutes (1981)およびThe Sambia (1986)は、母親や姉妹から引き離されて男子になるための過酷な成人式を何段階にわたり経験する過程を濃密に描き出すと同時に、妻との関係、母親との関係についても適度に目配りをしている。Sambia社会はcross-sex stateをことのほか抑圧する事例を提供しており、加えてsame-sex stateは出自集団を越えた地域の水準で強力に構築されている。しかしながらこうした例外的な状況にあっても、same-sex sateとcross-sex stateが相互に隠喩化される諸局面が明らかに認められるのであり、親族の進化が家族を消さなかったことについて、その受け入れを許すのである。
(1)人類学の歴史において、人類の進化は親族の形態という観点から論じられてきた。
(2)人類の「生存」「極限状況」に密接にかかわるテーマ―食糧確保・身体の防衛・生殖・社会化など―は、親族の働きをぬきにして語ることができない。
1.まずは、人類学における本研究の位置づけをおこなう。人類学の主要トピックであった親族は、分析と記述のための主要概念がE. LeachやR. Needhamによって次々に批判された1970年代以降、かつての勢いを失った。けれども90年代になると、考古学や霊長類学や認知諸科学の研究成果に突き動かされるかたちで、人類学者によるあたらしいタイプの親族研究が散見されるに至る。本発表はその動きと無縁でないが、あくまで従来の人類学を前提にしながら、二つの問題を設定する。
一つ目は家族の再評価であり、もう一つはジェンダーと贈与の節合の検討である。前者についていえば、従来の親族理論が「血族性と姻戚性」(consanguinity and affinity)を主軸として築かれており、核家族のような限定的モデルが軽視される傾向にあった点を背景として上げることができる。パプアニューギニアとマッシム地域の民族誌を読むかぎり、外婚的な出自集団は、一つの夫婦とその子どもを主要な成員とする家族を必ず併存させている。したがって、出自集団にとっての核家族の存在意義を明らかにする必要がある。後者については、M. StrathernのThe Gender of the Gift(1988)を参照せざるをえない。80年代以降、親族研究にとってかわったジェンダー研究に対し、人類学から大きな影響を与えた書物であり、核家族と出自集団に対しても深い洞察を得ることができるだろう。
これら二つの問題を、次のように言い換えてみよう。
(1)親族が血族と姻族を主軸にして成り立ち、二つの異なる血族集団間の女性のやりとりに注目するものだとしても、なぜそれによって一組のあたらしい夫婦とその家族をつくる必要があるのだろうか? 初期人類にプロト家族がすでに現れていたにせよ、集団間の縁組が進化する過程で、なお家族が生き残る必然はどこにあったのか?
(2)血族集団の内部に、また家族の内側にジェンダーは存在するが、贈与と結びつくことは基本的にはない。この二つは、ある集団が女性を外部の他の集団へと拠出するときに、婚資をはじめとする財の交換が集団間に生まれることによって、はじめて結びつくのである。贈与の原理が血族や家族にあっては支配的にならず、縁組と姻戚関係の創造において求められるということを、どのように理解すればよいのか?
2.Strathernがメラネシアにおける社会性として提起した二つの概念から出発して、考察を進めたい。same-sex stateおよびcross-sex stateという概念である。両者の内容は、およそ以下の対比によって示すことができる。
| same-sex state | cross-sex state |
| same-sex gender relations | cross-sex gender relations |
| political domain | domestic domain |
| mediated exchange | unmediated exchange |
| collective relations | particular relations |
| composite plurality | dual plurality |
メラネシア人の社会生活は家庭の領域でのcross-sex stateでないかぎり、家庭の外部におけるsame-sex stateとなり、そこでは男か女かのどちらかにジェンダー化された集合性を形成する。Strathernは両方の状況について、「人々は二つの実践を互いの隠喩になるように借用するので、実際には一つの実践がもう一つの形態の一部になっている」[1988:267]、と論じている。彼女は贈与儀礼のように特別な文脈において、same-sex stateがcross-sex stateへと類比されて現れることを強調する。より具体的には、次のように解釈できるだろう。贈与儀礼の成功に向けて、一つの出自集団が一つの家族のように協力しあうのである。男と女という二つの性別集団は、同一事業を営む夫婦のように無媒介的な交換、つまりは交換と呼べないほどの直接的なやりとりをつうじて、財を調達し儀礼の場を用意するわけである。
このようにcross-sex stateが 隠喩化したかたちでsame-sex stateに現れることは、一つの出自集団で生じるにしても、二つの別々の集団どうしに起こるとは思えない。というのも、このとき二つの集団は贈与の関係で結ばれるのであり、贈与の関係は家族というcross-sex stateにふさわしい原理ではないからである。そこで問題は、次の二つへと展開できる。
(1)cross-sex stateがsame-sex stateの内側へ隠喩として持ち込まれる状況についてわかるが、Strathernの指摘するように隠喩が相互に起こるとすれば、same-sex sateがcross-sex stateの内側へ隠喩化されるという逆の状況についても、明らかにする必要がある。これを明らかにできるならば、出自集団と家族は互いを必要としあい、親族の発達が家族を消さなかったことについても納得ができるだろう。
(2)ジェンダーの関係と贈与の原理の結びつきは、same-sex stateとcross-sex stateの相互に隠喩的な関係とどのようにつなげることが可能なのか。このつながりがわかれば、ジェンダーと贈与の必然的な関係を理解できるのではないか。
3.発表ではまず、後者の検討をおこなった。結論として、ジェンダーの領域と贈与の領域との間でも、相互的な隠喩の関係が生じていることを提起した。つまりは、縁組と贈与儀礼をともにおこなう二つの集団において、「人々は二つの実践を互いの隠喩になるように借用するので、実際には一つの実践がもう一つの形態の一部になっている」という状況を提示したのである。
縁組の出発点では、女を与える/受け取るという性に関する非対称が現れる。この非対称性への対処は、別の非対称性との類比をつうじておこなわれるのではないか。つまりは財の非対称性であり、贈与(代償)というかたちをとる、と考えることができる。ただし、類比は類比にすぎないので、ジェンダーと贈与は決して等価にはならないし、縁組で生じるジェンダーの非対称性が贈与によって対称性を回復することはない。女性の受け手(=夫方集団)は女性の与え手(=妻方集団)に対して、その非対称性を財の非対称性へと喩えてみせるだけである[父系制においてはこの非対称性に加えて、結婚で生まれる子どもの帰属についての非対称性が、さらに加わることになる]。
M. GodelierはThe Making of Great Men (1988)において、パプアニューギニア東高地の諸社会で実施される姉妹の交換婚を西高地での婚資と対比し、東高地では女性は女性とのみ等価なのだから、交換婚はモノの非対称性を導出しない、と論じている。しかしながら、東高地のさまざまな民族誌は、交換婚といえども受け手から与え手への贈与が絶えずおこなわれていることを報告する。交換婚は二つの結婚で構成されており、それぞれの結婚においてジェンダーと財との非対称性が生まれるのである。
このように考えると、ジェンダー関係と贈与―つまり財の非対称なやり取り―は、女性を外に出すときにはじめて結びつくことがわかる。つまり縁組によって、二つの非対称な関係が互いを参照し合うようになるのであり、結婚と贈与が不可分の結びつきを形成するわけである。
本発表は最後に(1)の問題の考察を、家族の詳細な記述を含む民族誌的な一事例に依拠して展開した。G. HerdtによるGuardians of the Flutes (1981)およびThe Sambia (1986)は、母親や姉妹から引き離されて男子になるための過酷な成人式を何段階にわたり経験する過程を濃密に描き出すと同時に、妻との関係、母親との関係についても適度に目配りをしている。Sambia社会はcross-sex stateをことのほか抑圧する事例を提供しており、加えてsame-sex stateは出自集団を越えた地域の水準で強力に構築されている。しかしながらこうした例外的な状況にあっても、same-sex sateとcross-sex stateが相互に隠喩化される諸局面が明らかに認められるのであり、親族の進化が家族を消さなかったことについて、その受け入れを許すのである。
2.死亡率:生活史としての理解と生態学としての理解(デイビッド・スプレイグ)
1.背景
人類進化の長い道筋を顧みて、ヒトの生活は持続可能(Sustainable)と考えてよいのであろうか。当然、現在までヒトは生存してきたことを見れば、ヒトの生活が持続していることは間違えないが、持続が保証されていたわけではなかったであろう。また、どのような生き物のとして生き延びてきたかを解明することは生態人類学の基本的な課題の一つである。
持続性を評価する上で最も基本的な要因の一つが死亡率といえる。持続することはすなわち生存することであり、生存を遮る死亡は生物界における基本現象であるとともに、その率は生物学理論の重要な説明要因でもある。進化論においては、自然選択の原動力であり、生態学では個体群の数の主要な決定要因である。近年の生物学では生き物の繁殖戦略が注目されがちなような気がするが、その対である死亡率について理論を整理しておく必要があると考えられる。死は、誕生とともに、生物がかならず経験する。繁殖は必ずしも全個体が経験するわけではない。むしろ殆どの生物個体は繁殖を経験することはない。何故ならば、繁殖の機会を得る前に死んでしまうからである。よって、繁殖戦略とともに、あるいはそれ以前に、死亡率を考慮して生物の進化と生態を評価する必要がある。
2.生態学と生活史理論における死亡率
一言で死亡率と言っても、生物学を構成する様々な理論のなかでその意味が一定ではないことに注意しなければならない。生物種の持続性を評価する理論体系は大きくいって生活史と生態学の視点、またはその組み合わせである。そこで、生活史と生態学の論旨のなかで果たす死亡率を整理しておく。
1)生態学(Ecology)
研究対象:生物の数と分布を説明しようとする研究分野。
研究する現象:研究対象となる現象は個体群の数と分布の変動と、それを引き起こす環境要因。
生態学における生活史の役割:生活史は前提条件として個体群変動モデル内で考慮するが、個体群変動によって生物の進化が伴う必要はない。
死亡率の役割:個体群変動の主要な要因であり、死亡率が高いと数が減少したり、分布が狭められたり、消滅したりする。
2)生活史(Life history)
研究対象:生物の生涯にわたる成長と繁殖の戦略を説明しようとする研究分野。
研究する現象:生涯の過程上の生活イベント(成長、成熟、繁殖、老化など)の連鎖とそのタイミング(順番や間隔)。
生活史理論における生態学の役割:生態学は生活史を説明する外部要因として扱われる場合が多く、生物の生活を左右する気候、生息地(森林・草地)、捕食者、環境の安定・不安定などが生活史の説明要因として扱われる。
死亡率の役割:生涯にわたる死亡率の「予定」は他の生活イベントの予定、特に繁殖の予定に影響する。生き物としての課題は死んでしまう前に子孫を残すこと。例えば、早死にする生物は早く繁殖し、長生きする生物は繰り返し繁殖する機会を得る。
生態と生活史は互いに外部要因・決定要因・前提要因として扱われる場合もあるが、知らぬ間に融合していることもあるので要注意: r/K selection。
3.ヒトはK戦略か、r戦略か?
生活史と生態を組み合わせた理論がr/K理論である。死亡率が低く、長生きして少数の子供を大事に育てる動物は生息地の生態的許容量(Carrying capacity, K)まで個体群を増やして、巧妙な生存戦略をもって安定的に生活する。逆に、死亡率が高く、寿命の短い動物は旺盛な繁殖力(Reproductive rate, r)と拡散力をもって生息環境の変動を乗り切る。生活史として、ヒトは、哺乳動物のなかでは大柄で、生存率も高く、子供が少ないので、通常はK戦略の動物と思われている。
ところが、そこで、K周辺まで増加した個体数を制御するはずの「密度効果」が問題になる。ヒトの人口を制御する密度効果のメカニズムは何なのか?黙示録の四騎士なのか?:悪疫 (pestilence)、戦争 (war)、飢饉 (famine)、死 (death)。このように考えると、K周辺の生活の実態は厳しく、生態学でいう死亡率は必ずしも低いとはいえない可能性がある。K周辺の生活の例に近世の日本をあげられる。近世の日本は巧妙な農林業戦略によって繁栄したともいえるが、同時に何度も大飢饉を体験し、近世後期はむしろ人口を減少させている。すなわち、持続可能な社会生態はある意味で全体として豊かでありながら、あらゆる密度効果のために生き物の実感としては厳しい生活かもしれない。
しかし、視点を変えるとヒトの生活史にはr戦略の側面もあると考えられる。まず、大型類人猿としては、ヒトの生存率のみならず繁殖率もかなり高い。ヒトは他の類人猿にはない「子供期」を生活史に組み込み、子供を早めに離乳させ、共同で子育てしながら、繰り返し出産することにより、繁殖力を向上させてきた。さらに、約20万年ものあいだ、南部アフリカから出発した人類は新天地を求めて全世界へと生息地を急拡大させ、人口は増加し続けてきた。また、度重なる飢饉や疫病による人口減少を乗り切ってきた。この見方では、ヒトは強烈なr戦略生物ともいえる。しかし、究極的にはr戦略は持続しにくいという前提がr戦略理論に潜んでいる。r戦略生物の運命は、自らがもたらす環境改変や密度効果により、増加路線の限界にぶつかる。そこで、新天地を求めてさらに拡散するか、K戦略へと移行せざるを得ない。
4.総括
人類進化の長い道筋を顧みて、ヒトの生活は持続可能(Sustainable)と考えてよいのであろうか。当然、現在までヒトは生存してきたことを見れば、ヒトの生活が持続していることは間違えないが、持続が保証されていたわけではなかったであろう。また、どのような生き物のとして生き延びてきたかを解明することは生態人類学の基本的な課題の一つである。
持続性を評価する上で最も基本的な要因の一つが死亡率といえる。持続することはすなわち生存することであり、生存を遮る死亡は生物界における基本現象であるとともに、その率は生物学理論の重要な説明要因でもある。進化論においては、自然選択の原動力であり、生態学では個体群の数の主要な決定要因である。近年の生物学では生き物の繁殖戦略が注目されがちなような気がするが、その対である死亡率について理論を整理しておく必要があると考えられる。死は、誕生とともに、生物がかならず経験する。繁殖は必ずしも全個体が経験するわけではない。むしろ殆どの生物個体は繁殖を経験することはない。何故ならば、繁殖の機会を得る前に死んでしまうからである。よって、繁殖戦略とともに、あるいはそれ以前に、死亡率を考慮して生物の進化と生態を評価する必要がある。
2.生態学と生活史理論における死亡率
一言で死亡率と言っても、生物学を構成する様々な理論のなかでその意味が一定ではないことに注意しなければならない。生物種の持続性を評価する理論体系は大きくいって生活史と生態学の視点、またはその組み合わせである。そこで、生活史と生態学の論旨のなかで果たす死亡率を整理しておく。
1)生態学(Ecology)
研究対象:生物の数と分布を説明しようとする研究分野。
研究する現象:研究対象となる現象は個体群の数と分布の変動と、それを引き起こす環境要因。
生態学における生活史の役割:生活史は前提条件として個体群変動モデル内で考慮するが、個体群変動によって生物の進化が伴う必要はない。
死亡率の役割:個体群変動の主要な要因であり、死亡率が高いと数が減少したり、分布が狭められたり、消滅したりする。
2)生活史(Life history)
研究対象:生物の生涯にわたる成長と繁殖の戦略を説明しようとする研究分野。
研究する現象:生涯の過程上の生活イベント(成長、成熟、繁殖、老化など)の連鎖とそのタイミング(順番や間隔)。
生活史理論における生態学の役割:生態学は生活史を説明する外部要因として扱われる場合が多く、生物の生活を左右する気候、生息地(森林・草地)、捕食者、環境の安定・不安定などが生活史の説明要因として扱われる。
死亡率の役割:生涯にわたる死亡率の「予定」は他の生活イベントの予定、特に繁殖の予定に影響する。生き物としての課題は死んでしまう前に子孫を残すこと。例えば、早死にする生物は早く繁殖し、長生きする生物は繰り返し繁殖する機会を得る。
生態と生活史は互いに外部要因・決定要因・前提要因として扱われる場合もあるが、知らぬ間に融合していることもあるので要注意: r/K selection。
3.ヒトはK戦略か、r戦略か?
生活史と生態を組み合わせた理論がr/K理論である。死亡率が低く、長生きして少数の子供を大事に育てる動物は生息地の生態的許容量(Carrying capacity, K)まで個体群を増やして、巧妙な生存戦略をもって安定的に生活する。逆に、死亡率が高く、寿命の短い動物は旺盛な繁殖力(Reproductive rate, r)と拡散力をもって生息環境の変動を乗り切る。生活史として、ヒトは、哺乳動物のなかでは大柄で、生存率も高く、子供が少ないので、通常はK戦略の動物と思われている。
ところが、そこで、K周辺まで増加した個体数を制御するはずの「密度効果」が問題になる。ヒトの人口を制御する密度効果のメカニズムは何なのか?黙示録の四騎士なのか?:悪疫 (pestilence)、戦争 (war)、飢饉 (famine)、死 (death)。このように考えると、K周辺の生活の実態は厳しく、生態学でいう死亡率は必ずしも低いとはいえない可能性がある。K周辺の生活の例に近世の日本をあげられる。近世の日本は巧妙な農林業戦略によって繁栄したともいえるが、同時に何度も大飢饉を体験し、近世後期はむしろ人口を減少させている。すなわち、持続可能な社会生態はある意味で全体として豊かでありながら、あらゆる密度効果のために生き物の実感としては厳しい生活かもしれない。
しかし、視点を変えるとヒトの生活史にはr戦略の側面もあると考えられる。まず、大型類人猿としては、ヒトの生存率のみならず繁殖率もかなり高い。ヒトは他の類人猿にはない「子供期」を生活史に組み込み、子供を早めに離乳させ、共同で子育てしながら、繰り返し出産することにより、繁殖力を向上させてきた。さらに、約20万年ものあいだ、南部アフリカから出発した人類は新天地を求めて全世界へと生息地を急拡大させ、人口は増加し続けてきた。また、度重なる飢饉や疫病による人口減少を乗り切ってきた。この見方では、ヒトは強烈なr戦略生物ともいえる。しかし、究極的にはr戦略は持続しにくいという前提がr戦略理論に潜んでいる。r戦略生物の運命は、自らがもたらす環境改変や密度効果により、増加路線の限界にぶつかる。そこで、新天地を求めてさらに拡散するか、K戦略へと移行せざるを得ない。
4.総括
- 生活の持続に失敗すれば死んでしまう。コミュにティーが縮小したり消滅したりする。ただし、高い死亡率が継続されると生活史が進化する可能性もある。
- K近くの生活は厳しい。生息環境が「豊か」であっても、密度効果のために生物の実感は厳しい。「持続的」な社会は非常に厳しい社会かもしれない。
- r戦略はとりあえず生物の実感としては豊かな生活かもしれない。けれど時限付き。
- 個体群が成長し続けるには新天地を求めるか、何らかの技術革新(ヒトの場合は道具使用や生業の発展)でKを上げなければならない。
- ヒトはr/K戦略を使い分けてきたのではないか。技術と組織で生存率・繁殖率を高く維持してきた。ただし、生活史の進化はなぜか主に出産間隔を縮めることで対応?離乳食という技術?
- 現代の少子化は貨幣経済を媒介とした密度効果の表れ?
3.ホロコーストの生存者たちとともに生きる人びと:アウシュヴィッツ=ビルケナウ強制収容所のガイドたち(田中雅一)
死者だけでなく、多くの生存者survivorや犠牲者victimを生み出す災害や戦争の記憶をいかに継承すべきなのか。生存者らの悲痛な経験をいかに共有できるのか。こうした問題意識は、災害や戦争に関わってきた歴史家や医療関係者、司法関係者、芸術家、さらにわたしたち文化人類学者に共通して認められる。本発表では、災害地や戦場など、いわゆる人類の負の遺産を案内するガイドを取り上げて、その性格を考えてみたい。具体的には、2015年に170万人が訪れたポーランド・アウシュヴィッツで働くガイドたちを取り上げる。
ガイドたちは、訪問者が集中する夏には毎日2回アウシュヴィッツ博物館に展示されている遺品を訪問者たちに紹介している。遺品の中にはユダヤ人たちが残したスーツケースやおびただしい数の靴、義足などが含まれている。また、髪の毛で編まれた絨毯も展示されている。仕事とはいえ、こうした遺品に毎日対面するには、かなりの精神力が必要である。
心身への影響を恐れ、展示品にあまりにも無関心になろうとすると、訪問者への説明が機械的になってアウシュヴィッツのできごとを十分に伝えることができない。これは、程度の差はあれ、アウシュヴィッツに限らず、広島や長崎、沖縄戦のガイドにも当てはまるであろう。つまり、負の遺産のガイドは複雑な「感情労働」に従事していると考えられる。ここで言う感情労働とは、社会学者ホックシールドが『管理される心』(1983年)に提案した概念で、「自身の感情表現を管理し〔たとえば微笑みを絶やさない、配慮の行き届いたしぐさ〕、サービスを受ける客に肯定的な感情〔安心感〕を生みだしたり、否定的な感情〔不安や恐怖〕を軽減したりする一連の行為」を意味する。日本では、主として介護や看護、保育の分野で研究がなされてきたが、ツーリスト・ガイドについての研究は皆無である。
本発表では、以上のような問題意識を念頭に、4人の女性ガイドと1人の男性ガイドの声を紹介する。ガイドたちは、アウシュヴィッツには負のエネルギーや負の感情が認められると述べる。また、展示品自体もしばしば恐怖を引き起こす。一人のガイドは、始めて2年間は陳列ケースに展示されている女性の髪を直視できなかったと語っている。このような展示の延長に生存者の存在を位置づけることも可能である。展示品は、いつも同じところに同じものが同じ様態で展示されている。このため、最初は衝撃的であっても、すこしずつ慣れてくる。しかし、生存者は違う。いつどんな生存者に会うのか予想がつかないし、どんなことを話し始めるのかも分からない。あるガイドは、ホロコーストとある程度距離をとって接していたが生存者と接すると、そのような距離が攪乱し、自分の生活に入ってくる。そして、気分が悪くなったり、夢に生存者が現れたりするという。もちろん、生存者に接することで、ガイドの語りはより豊かになり真正性を増す。この真正性がないと、ガイドの語りから過去のできごとへの想像力が働かない。しかし、否定的な側面も無視できない。
訪問者との関係では、ガイドは感情を込めて案内する必要がある。そうでないと本物として受け止められない。しかし、自分の感情を前面に出しすぎると、ガイドが中心になってしまう。ガイドは俳優ではない。あくまで媒介者でなければならない。このため、あるガイドは、感情を抑え、説明をすることに徹する。これら二つは、感情についての対立する態度と言える。まったく感情を配すると嘘っぽくなり、反対に感情を出しすぎると過去のできごとが背景に退いてしまう。なんらかのバランスが必要である。
アウシュヴィッツで感じる悪い感情を克服するために、ガイドたちはさまざまな工夫をこらしている。博物館にはガイドの精神衛生を扱う専門家はいない。このため、ガイドたちは親しい同僚と話し合う機会を設けたり、家庭と仕事をできるだけ分離するなどして、対処している。気晴らしにどこかに行ったり、ホロコーストと関係のない読書に熱中したりする。しかし、耐えられずに競争率の高いガイドの道を諦める人もいる。
ガイドたちは、おおよそ仕事に深くコミットしていて、勉強熱心である。Missionとpassionが不可欠だと述べる。あるガイドは、その仕事を(生徒が毎日変わる)教師の役割にたとえている。
すでに指摘したように、感情の統御が重要であることが明らかになった。これは、冒頭で示唆したようにアウシュヴィッツに限らず、負の遺産のガイド一般に言えることであろう。言い換えると、日本では福島などをダーク・ツーリズムの拠点にすることで、経済的な復興を目指そうという動きがあるが、これを実現するためには、ガイドをはじめとする関係者の「心のケア」を無視すべきではない。
本発表では、ほかにイスラエルからの訪問者、ドイツからの訪問者に対するガイドの態度、接する際に気をつけなければならないこと、アウシュヴィッツにおける「カトリック化」をめぐる問題などに触れた。
ガイドたちは、訪問者が集中する夏には毎日2回アウシュヴィッツ博物館に展示されている遺品を訪問者たちに紹介している。遺品の中にはユダヤ人たちが残したスーツケースやおびただしい数の靴、義足などが含まれている。また、髪の毛で編まれた絨毯も展示されている。仕事とはいえ、こうした遺品に毎日対面するには、かなりの精神力が必要である。
心身への影響を恐れ、展示品にあまりにも無関心になろうとすると、訪問者への説明が機械的になってアウシュヴィッツのできごとを十分に伝えることができない。これは、程度の差はあれ、アウシュヴィッツに限らず、広島や長崎、沖縄戦のガイドにも当てはまるであろう。つまり、負の遺産のガイドは複雑な「感情労働」に従事していると考えられる。ここで言う感情労働とは、社会学者ホックシールドが『管理される心』(1983年)に提案した概念で、「自身の感情表現を管理し〔たとえば微笑みを絶やさない、配慮の行き届いたしぐさ〕、サービスを受ける客に肯定的な感情〔安心感〕を生みだしたり、否定的な感情〔不安や恐怖〕を軽減したりする一連の行為」を意味する。日本では、主として介護や看護、保育の分野で研究がなされてきたが、ツーリスト・ガイドについての研究は皆無である。
本発表では、以上のような問題意識を念頭に、4人の女性ガイドと1人の男性ガイドの声を紹介する。ガイドたちは、アウシュヴィッツには負のエネルギーや負の感情が認められると述べる。また、展示品自体もしばしば恐怖を引き起こす。一人のガイドは、始めて2年間は陳列ケースに展示されている女性の髪を直視できなかったと語っている。このような展示の延長に生存者の存在を位置づけることも可能である。展示品は、いつも同じところに同じものが同じ様態で展示されている。このため、最初は衝撃的であっても、すこしずつ慣れてくる。しかし、生存者は違う。いつどんな生存者に会うのか予想がつかないし、どんなことを話し始めるのかも分からない。あるガイドは、ホロコーストとある程度距離をとって接していたが生存者と接すると、そのような距離が攪乱し、自分の生活に入ってくる。そして、気分が悪くなったり、夢に生存者が現れたりするという。もちろん、生存者に接することで、ガイドの語りはより豊かになり真正性を増す。この真正性がないと、ガイドの語りから過去のできごとへの想像力が働かない。しかし、否定的な側面も無視できない。
訪問者との関係では、ガイドは感情を込めて案内する必要がある。そうでないと本物として受け止められない。しかし、自分の感情を前面に出しすぎると、ガイドが中心になってしまう。ガイドは俳優ではない。あくまで媒介者でなければならない。このため、あるガイドは、感情を抑え、説明をすることに徹する。これら二つは、感情についての対立する態度と言える。まったく感情を配すると嘘っぽくなり、反対に感情を出しすぎると過去のできごとが背景に退いてしまう。なんらかのバランスが必要である。
アウシュヴィッツで感じる悪い感情を克服するために、ガイドたちはさまざまな工夫をこらしている。博物館にはガイドの精神衛生を扱う専門家はいない。このため、ガイドたちは親しい同僚と話し合う機会を設けたり、家庭と仕事をできるだけ分離するなどして、対処している。気晴らしにどこかに行ったり、ホロコーストと関係のない読書に熱中したりする。しかし、耐えられずに競争率の高いガイドの道を諦める人もいる。
ガイドたちは、おおよそ仕事に深くコミットしていて、勉強熱心である。Missionとpassionが不可欠だと述べる。あるガイドは、その仕事を(生徒が毎日変わる)教師の役割にたとえている。
すでに指摘したように、感情の統御が重要であることが明らかになった。これは、冒頭で示唆したようにアウシュヴィッツに限らず、負の遺産のガイド一般に言えることであろう。言い換えると、日本では福島などをダーク・ツーリズムの拠点にすることで、経済的な復興を目指そうという動きがあるが、これを実現するためには、ガイドをはじめとする関係者の「心のケア」を無視すべきではない。
本発表では、ほかにイスラエルからの訪問者、ドイツからの訪問者に対するガイドの態度、接する際に気をつけなければならないこと、アウシュヴィッツにおける「カトリック化」をめぐる問題などに触れた。
4.相互行為システムによる二重の層位での選択―「地域社会の消滅」という課題に向き合うときに見えてくるもの―(北村光二)
1.はじめに
この報告では、相互行為システムのコミュニケーションが卓越するような社会におけるコミュニケーション現象がどのような特徴をもつものであるかを検討する。ここでいう「相互行為システムのコミュニケーション」とは、個々人が自律的に外界との直接的な関係を作り出そうとすることを前提に行う仲間とのコミュニケーションであり、基本的には、参与する人々が同じ場所に居合わせて相互に知覚し合うことで可能になっているのである。そしてそれは、あらかじめ存在する役割関係や権利・義務関係を用いて集団的意思決定を行い、それにもとづく集団的問題対処を行う「組織システムのコミュニケーション」とは明確に区別できるものである。さらに、それは、参与者の一方が拒否するだけで存在することをやめてしまう状況システムでもあるという点も重要である。
以下での考察の出発点は、私が1986年以来調査を継続してきた北ケニア牧畜民トゥルカナで見出され、私が「強要的な物乞い」と名付けた相互行為である。そのコミュニケーション現象をどう理解したらよいかについて考察を進めながら、それをより高い抽象度の用語によって記述することによって、この現象がトゥルカナに特別な、ないしは、制度や権威構造が未発達な原初的な社会に特別なものではないことを明らかにし、これに類似のより多様な現象を理解可能にする理論的枠組みについての新たな提案を行いたい。そして最後に、この枠組みを用いて、現代日本が抱える「地域社会の消滅」という問題にもアプローチする。
2.トゥルカナに特徴的なコミュニケーションのあり方をどう理解するか
私は、長年にわたるトゥルカナとのつき合いの中で、近代に生きる私たちにとって強烈な印象を刻むものとなる「強要的な物乞い」という相互行為について、トゥルカナ自身がそれをどのようなコミュニケーションとして体験しているのかを明らかにしようと悪戦苦闘してきた。彼らは、さまざまな場面を捉えて相手に「もの」を与えるように要求するのだが、相手も要求されるごとに与えることなどできないわけで、それを断ろうとすることでそこに交渉的な相互行為が繰り広げられることになる.この相互行為についての理解を困難にしているものとは、第一に、物を乞うて相手が拒否したとしても乞う側がその獲得をあきらめないために、ないしは、乞われる側が拒否してもそのやりとりを打ち切ることができないために、その要求と拒否というやりとりが終わりのないもののようにいつまでも続くという点であり、第二には、交渉の当事者として、このような膠着状態の先にどのような決着の可能性があり得ると想定しているのかが理解不能だという点である。
ここでトゥルカナが何をしているのかについてのとりあえずの理解は以下のようになる。まず、双方の当事者があくまでも各自の利益を確保しようとすることによって膠着状態に至ることになるが、そのまま当事者間の交渉を継続するというやり方を貫くことによってそのような事態に何とか対処しようとしているのだが、同時に、あくまでも当事者たちが一致して妥当だと判断することになる対処法を選択しようともしているのである。そのうえで、ここで対処法として何を選択することになるのかはそのときの相互行為の行きがかりに委ねられることになるのであるが、この点も私たちの理解を難しくしているこのやり方に特徴的な部分である。
そのような振舞い方を支えている彼らの基本的な選択とは以下のようなものだと考えられる。まず、彼らは、そのときの相互行為によって直面する課題に関わる何らかの結果を実現しようとするとともに、個々の相互行為を超え出るより大きな地域社会の秩序に関わる選択として、仲間との共存を再生産可能のものとして維持しようともしているのだと考えなければならない。その際、それぞれの当事者はその選択を、自分自身の選択としても経験しながら、そのときの相互行為の相手とともにする「われわれの選択」としても経験しているのである。そしてこのようなより大きな地域社会に関わる選択は、現代日本に生きる私たちにはごく特別なものに見えるのであり、私たちは私的な領域における選択の自由を手にした一方で、より大きな社会の秩序に関わる選択に仲間とともに当事者として直接関与するという経験を喪失してしまったかのようなのである。
しかし、そのような感慨は人類学者が陥りがちなノスタルジアに過ぎないともいえるはずであり、彼らがしていることをごく特別なこととして祭り上げてしまうことなく、人間であれば誰もが当たり前にしているはずのことの一表現型として理解する道を探さなければならないはずである。そのような考え方を手にすることによって、民族誌的研究にもとづく新たな人間理解の可能性が切り開かれるはずであるのだから。
2.新しい理解
なぜ、彼らのやり方が特別なものに見えるのかを考えるところから始めよう。まず、彼らが徹底的に個人の利益の確保にこだわりながら、同時に仲間との協働によって、仲間と共有される対処法を選択しようとしているという点が重要である。そして、最終的に何らかの決着がもたらされたときには、たんなる個人の選択にすぎなかったはずものが、仲間との協働による「われわれの選択」になっていると考えられるのである。
その選択は、何らかの個別的な結果を志向するものであるとともに、その選択の前提となる人びとが暮らす地域社会の秩序の再生産を志向するものでもあるという、二重の層位にわたる選択になっているのである。そして、このような性質を持つ選択は、近代に生きる私たちには全くなじみのないものだというのではない。私たちの場合は、例えば、直面する問題に仲間と協働して対処しようとして、人びとが共有する規則に従った行為選択を行うというときにしていることなのである。それは、個別的な問題対処を志向するものであると同時に、社会の秩序維持を志向するものでもあるのだから。ただしこの場合は、問題への対処法として何が選択されることになるかは規則によってあらかじめ指定されているのに対して、彼らのやり方では、それが何になるかはそのときの相互行為の行きがかりに委ねられているのである。
規則に従った仲間との協働による問題対処の場合を参照すれば明らかなように、このような選択は、決して、個人の、すなわち、意識システムの選択ではなく、それとは明確に区別できる社会システムの選択である。そして、トゥルカナの場合には、「相互行為システムの選択」を問題にしなければならない。さらに、その選択はそのときの相互行為の行きがかりに委ねられているという意味で、相互行為システムの主体的な選択だと考えられなければならない。個人がする主体的な選択とは、そのときの具体的レベルでの選択に先立って、その前提となる可能な選択肢集合の全体を選択した上でなされるのであり、それは二重の層位での選択ということになる。選択肢として他なる可能性があることを知ったうえで、あえてその他なる可能性を実現しなかったことによって生起しているものという意味で、この場合は相互行為システムの主体的な選択だということになる。
3.オートポイエーシスを確実にするメタレベルの選択
ここで問題にしている相互行為システムがする二重の層位での選択とは、直面する問題への対処のためにする具体的レベルの選択において、その操作それ自体についての反省が求められるところで、その操作の前提となる、個々の相互行為を超え出る地域社会の秩序をある望ましいものとして確保しようとする選択が顕在化したものだとも理解できる。そこで確保しようとしている秩序とは、基本的には、コミュニケーションの試みにコミュニケーションの試みが接続して相互行為が継続することをより容易にするような状態のことであり、コミュニケーションのオートポイエーシス(=自己産出)を背後から支える状態のことである。具体的には、この相互行為システムのコミュニケーションにおいて、それぞれの当事者が、相手の自律的な選択を尊重して、たとえそれが自らの期待に反するものであってもコミュニケーションの接続を放棄せずに、あくまでもそこに共通理解を見出そうとし続けるというやり方を採用することによって、コミュニケーションの接続を容易で望ましいものにするような相互行為の場がそこに確保されることになるのだと考えられるのである。そして、このときのより大きな社会の秩序に関わる選択とは、具体的レベルの選択に随伴するメタレベルの選択という位置づけになるはずである。
私たちも、コミュニケーションの安定的再生産を確保しようとして、相手の意向に合わせて交渉をまとめようとすることがあるが、それは個々の相互行為を超え出る社会が選択してしまっている秩序のあり方(と当事者が考えているもの)に従属することにしかならない。それは当事者個人の主体的な選択ではないというだけではなく、そのときの相互行為システムの選択にさえなっていない。具体的には、例えば、当事者相互の利害が対立する問題への対処として私たちが採用するふつうのやり方とは、規則や制度のような社会が選択した基準に従って、そのときの利害の対立を調停するような妥当なプランについての合意を形成しようとするというものである。そして、解決が難しい課題に仲間と協働して対処しようとするときに私たちが採用するふつうのやり方とは、そのときのコミュニケーションに先立って存在している「組織」を前提にした集団的意思決定によるものであり、その場合には組織のすべてのメンバーがその決定に従属せざるを得なくなるというものになる。これらのものは、同じ場所に居合わせてコミュニケーションしている人々の主体的な参与による相互行為システムの選択とは、明らかに別のものになる。
その一方で、集団的意思決定を容易にする「組織」を持たず、規則や制度のような社会という個々の相互行為を超え出るまとまりが用意する支えが未発達な、人間以前の社会や人間の原初的な社会では、相互行為システムにおける二重の選択という現象は、ごく一般的なものである。哺乳類の群居的な社会においては、それぞれの個体が一般的な行動傾向として、近くにいる同種個体と活動を同調させようとしながら可能な限りコンフリクトを回避しようとするというやり方を採用することで、そのような状態が繰り返し再生産されるという社会秩序が確保され、それによって同種個体間でのこの種の行為接続が、個別的な出来事として容易に選択できるようになっている。真猿類の群れ社会では、同じ群れのメンバーどうしという関係で活動の同調とコンフリクトの回避を行うことによって、群れというまとまりが維持されてコミュニケーションの安定的再生産を保証する秩序が成立し、その秩序のもとで同じ群れのメンバー間での様々なタイプの行為接続がより容易に選択できるようになっているのである。
チンパンジー属の社会では、同じ集団の他のメンバーと離れて暮らせるようになっているが、離れて暮らしていた個体間で同じ集団のメンバーどうしという関係にあることを確認する手段と機会が用意されていることから、それらを活用することによって同じ集団のメンバーどうしのコミュニケーションの安定的再生産を保証する秩序が成立しているのだと考えられる。さらに、相互行為システムのコミュニケーションが卓越する原初的な人間社会においても、例えば、狩猟採集民ブッシュマンの社会でも、トゥルカナ同様に、徹底的に個人の自律性を尊重し合うというやり方でコミュニケーションの安定的再生産を確保しようとするメタレベルの選択が、不可欠のものとして実行されている。
すでに述べたように、現代日本に生きる私たちにとっては、相互行為システムのコミュニケーションにおいて自らが当事者として、個々の相互行為を超え出るより大きな社会の秩序に関わる選択に直接関与するという経験は、ごく特別なものになってしまっているのだと思える。しかし、以下に示すような相互行為システムのコミュニケーションが重要な機能を果たしている地域社会においては、トゥルカナ社会において私が見出したものと類似の現象が成立しているのではないかと思える。具体的に検討してみよう。
4.島社会における「島で暮らし続ける」という選択
現代日本では、生まれ育った島を出て本土で暮らすということが少しも特別な選択ではなくなってきている。そのような状況では、そこで暮らし続けようとすることは、より積極的な選択として自ら肯定できるものにならなければならなくなっている。そして、「島で暮らし続ける」という選択とは、そのように選択した者にとっては、たんに個人として島から出ないというだけのことではなく、この島の地域社会をこれから先も存続し続けるものにしようとするという覚悟のもとでそうすることになるのだと考えられるのである。それは、「地域社会の消滅」という最悪の事態を起こりうる未来として自覚したうえで、日々の問題対処のための相互行為を実行しながら、同時に、そのときの選択が個々の相互行為を超え出る地域的なコミュニケーションのまとまりの再生産をより容易にするものになっているかを繰り返し反省するようになるということである。
以下では、以前に私が調査した岡山県白石島の事例を手がかりに考察を進めよう。その研究では島社会が抱えるさまざまな問題を取り上げたが、ここでは地域の伝統芸能である「白石踊り」の継承の巡る問題に焦点を当てて検討する。
この白石踊りは、かつては島民のほとんどが踊れた地域の盆踊りであったが、1976年に国の重要無形民俗文化財に指定され、近年は観光の目玉とも目されており、「白石島の誇り」として島民に意識されてもいる。しかし、過疎化による人口減少と、娯楽の多様化による島民の踊り離れもあって、島民のための盆踊りの輪は小規模化し、以前のような盛り上がりは期待すべくもないものになっている。このような状態はかなり以前から誰の目にも明らかなものになっており、伝統行事継承への危機感から、小中学校で子供たちに踊りを教えるという試みが20年以上も続けられている。また、お盆が近づく時期には、観光客向けに踊りの機会を設けたり、チャーター船を仕立てた鑑賞体験ツアーを企画して、一緒に踊りを体験できるようにレクチャーする機会を提供してもいる。
このような伝統行事継承のための活動は、「白石踊会」という島民の中でもとくに熱心な人たちによって担われているが、その活動に対する他の島民の反応は、予想外に懐疑的なものなのである。観光客向けの踊りでは、揃いの衣装で見栄えの良いものが披露されるとともに、踊りの最中に解説が挿入されて中断したりするが、それに対して「これでは誰のための白石踊りかわからない」という不満が述べられる。また、学校教育の場での踊りの指導に対しても、島民にとっての踊りのあるべき姿という観点から、「否応なく習うものになった」という否定的な評価が述べられたりもする。文化財指定という肩書に対してさえ、「文化財に指定されたから伝承しているのではない」という島の人びとの「自分たちの踊り」への誇りが述べられたりもするのである。
しかし、だからといって、外部からの高い評価や経済的効果それ自体を否定的に考えているわけではない。それらは、地元の年中行事として自分たちが大切にしてきたという思いと共に、継承の危機に際してそのあるべき姿を意識することを動機づける重要な要素になっているのである。外部からの要請にも応えながら、その行事のあるべき姿との間に折り合いをつけようと試行錯誤を繰り返してきているのである。このようなやり方で白石島の人びとが実現しようとしていることとは、この行事を今後とも長期間にわたって維持することを可能にするような地域社会の秩序を確保することなのだと考えられる。この場合人びとは、この行事に対する個人的な好みの違いから誰もが同じように積極的に関与するとは限らないということを前提に、それぞれがそれぞれの立場でこの問題に取り組もうとしたときに、それを多くの島民が納得するものへとまとめ上げようとコミュニケーションを接続し続けることが不可欠となるが、それをより容易にするような秩序を確保しようとしているということなのである。
ここで白石島の人びとがしていることは、相互行為システムがする選択という操作それ自体について反省することによって、その前提となる、個々の相互行為を超え出る地域社会の秩序をある望ましいものとして確保しようとすることそのものだと考えられる。
5.おわりに
地域的な広がりの内部におけるコミュニケーションの接続をより容易なものとして確保しようするメタレベルの選択とは、個人の自律的な選択と矛盾しない、共に暮らす仲間との協働による「われわれの選択」によって、すなわち、相互行為システムの主体的な選択によって自らの人生を切り開こうとする生き方にとって、必要不可欠なものになると考えられるのである。ただし、近代に生きる私たちの場合は、仲間との協働による集団的な問題対処において、外部的な権威や制度が指定するものに従属することで仲間との協働が可能になっていることがふつうなのである。その場合には、「相互行為システムの主体的な選択」が問題になることはなく、この種のメタレベルの選択に目が向けられることもない。
ここで注目したようなメタレベルの選択が顕在化することは、相互行為システムのコミュニケーションが当事者の一方が拒否するだけで存在することをやめてしまう状況システムであるという事実に直接由来している。コミュニケーションの地域的まとまりを強化する制度的支えや権威構造を欠いている自然発生的な地域社会では、相互行為システムのコミュニケーションが卓越する一方で、そこでのコミュニケーションの接続が簡単に途絶しないようにしようとして、この種の選択が不可避的に顕在化するのだと考えられる。また、現代日本の島社会をはじめとする消滅可能性が取りざたされる地域社会では、従来からあった組織的・制度的支えの弱体化に伴い、相互行為システムのコミュニケーションの重要性が無視できないものになるのであり、地域的な広がりにおけるこの種のコミュニケーションの接続を確保しようとする選択がより強調された形で顕在化することになるのだと考えられるはずである。
この報告では、相互行為システムのコミュニケーションが卓越するような社会におけるコミュニケーション現象がどのような特徴をもつものであるかを検討する。ここでいう「相互行為システムのコミュニケーション」とは、個々人が自律的に外界との直接的な関係を作り出そうとすることを前提に行う仲間とのコミュニケーションであり、基本的には、参与する人々が同じ場所に居合わせて相互に知覚し合うことで可能になっているのである。そしてそれは、あらかじめ存在する役割関係や権利・義務関係を用いて集団的意思決定を行い、それにもとづく集団的問題対処を行う「組織システムのコミュニケーション」とは明確に区別できるものである。さらに、それは、参与者の一方が拒否するだけで存在することをやめてしまう状況システムでもあるという点も重要である。
以下での考察の出発点は、私が1986年以来調査を継続してきた北ケニア牧畜民トゥルカナで見出され、私が「強要的な物乞い」と名付けた相互行為である。そのコミュニケーション現象をどう理解したらよいかについて考察を進めながら、それをより高い抽象度の用語によって記述することによって、この現象がトゥルカナに特別な、ないしは、制度や権威構造が未発達な原初的な社会に特別なものではないことを明らかにし、これに類似のより多様な現象を理解可能にする理論的枠組みについての新たな提案を行いたい。そして最後に、この枠組みを用いて、現代日本が抱える「地域社会の消滅」という問題にもアプローチする。
2.トゥルカナに特徴的なコミュニケーションのあり方をどう理解するか
私は、長年にわたるトゥルカナとのつき合いの中で、近代に生きる私たちにとって強烈な印象を刻むものとなる「強要的な物乞い」という相互行為について、トゥルカナ自身がそれをどのようなコミュニケーションとして体験しているのかを明らかにしようと悪戦苦闘してきた。彼らは、さまざまな場面を捉えて相手に「もの」を与えるように要求するのだが、相手も要求されるごとに与えることなどできないわけで、それを断ろうとすることでそこに交渉的な相互行為が繰り広げられることになる.この相互行為についての理解を困難にしているものとは、第一に、物を乞うて相手が拒否したとしても乞う側がその獲得をあきらめないために、ないしは、乞われる側が拒否してもそのやりとりを打ち切ることができないために、その要求と拒否というやりとりが終わりのないもののようにいつまでも続くという点であり、第二には、交渉の当事者として、このような膠着状態の先にどのような決着の可能性があり得ると想定しているのかが理解不能だという点である。
ここでトゥルカナが何をしているのかについてのとりあえずの理解は以下のようになる。まず、双方の当事者があくまでも各自の利益を確保しようとすることによって膠着状態に至ることになるが、そのまま当事者間の交渉を継続するというやり方を貫くことによってそのような事態に何とか対処しようとしているのだが、同時に、あくまでも当事者たちが一致して妥当だと判断することになる対処法を選択しようともしているのである。そのうえで、ここで対処法として何を選択することになるのかはそのときの相互行為の行きがかりに委ねられることになるのであるが、この点も私たちの理解を難しくしているこのやり方に特徴的な部分である。
そのような振舞い方を支えている彼らの基本的な選択とは以下のようなものだと考えられる。まず、彼らは、そのときの相互行為によって直面する課題に関わる何らかの結果を実現しようとするとともに、個々の相互行為を超え出るより大きな地域社会の秩序に関わる選択として、仲間との共存を再生産可能のものとして維持しようともしているのだと考えなければならない。その際、それぞれの当事者はその選択を、自分自身の選択としても経験しながら、そのときの相互行為の相手とともにする「われわれの選択」としても経験しているのである。そしてこのようなより大きな地域社会に関わる選択は、現代日本に生きる私たちにはごく特別なものに見えるのであり、私たちは私的な領域における選択の自由を手にした一方で、より大きな社会の秩序に関わる選択に仲間とともに当事者として直接関与するという経験を喪失してしまったかのようなのである。
しかし、そのような感慨は人類学者が陥りがちなノスタルジアに過ぎないともいえるはずであり、彼らがしていることをごく特別なこととして祭り上げてしまうことなく、人間であれば誰もが当たり前にしているはずのことの一表現型として理解する道を探さなければならないはずである。そのような考え方を手にすることによって、民族誌的研究にもとづく新たな人間理解の可能性が切り開かれるはずであるのだから。
2.新しい理解
なぜ、彼らのやり方が特別なものに見えるのかを考えるところから始めよう。まず、彼らが徹底的に個人の利益の確保にこだわりながら、同時に仲間との協働によって、仲間と共有される対処法を選択しようとしているという点が重要である。そして、最終的に何らかの決着がもたらされたときには、たんなる個人の選択にすぎなかったはずものが、仲間との協働による「われわれの選択」になっていると考えられるのである。
その選択は、何らかの個別的な結果を志向するものであるとともに、その選択の前提となる人びとが暮らす地域社会の秩序の再生産を志向するものでもあるという、二重の層位にわたる選択になっているのである。そして、このような性質を持つ選択は、近代に生きる私たちには全くなじみのないものだというのではない。私たちの場合は、例えば、直面する問題に仲間と協働して対処しようとして、人びとが共有する規則に従った行為選択を行うというときにしていることなのである。それは、個別的な問題対処を志向するものであると同時に、社会の秩序維持を志向するものでもあるのだから。ただしこの場合は、問題への対処法として何が選択されることになるかは規則によってあらかじめ指定されているのに対して、彼らのやり方では、それが何になるかはそのときの相互行為の行きがかりに委ねられているのである。
規則に従った仲間との協働による問題対処の場合を参照すれば明らかなように、このような選択は、決して、個人の、すなわち、意識システムの選択ではなく、それとは明確に区別できる社会システムの選択である。そして、トゥルカナの場合には、「相互行為システムの選択」を問題にしなければならない。さらに、その選択はそのときの相互行為の行きがかりに委ねられているという意味で、相互行為システムの主体的な選択だと考えられなければならない。個人がする主体的な選択とは、そのときの具体的レベルでの選択に先立って、その前提となる可能な選択肢集合の全体を選択した上でなされるのであり、それは二重の層位での選択ということになる。選択肢として他なる可能性があることを知ったうえで、あえてその他なる可能性を実現しなかったことによって生起しているものという意味で、この場合は相互行為システムの主体的な選択だということになる。
3.オートポイエーシスを確実にするメタレベルの選択
ここで問題にしている相互行為システムがする二重の層位での選択とは、直面する問題への対処のためにする具体的レベルの選択において、その操作それ自体についての反省が求められるところで、その操作の前提となる、個々の相互行為を超え出る地域社会の秩序をある望ましいものとして確保しようとする選択が顕在化したものだとも理解できる。そこで確保しようとしている秩序とは、基本的には、コミュニケーションの試みにコミュニケーションの試みが接続して相互行為が継続することをより容易にするような状態のことであり、コミュニケーションのオートポイエーシス(=自己産出)を背後から支える状態のことである。具体的には、この相互行為システムのコミュニケーションにおいて、それぞれの当事者が、相手の自律的な選択を尊重して、たとえそれが自らの期待に反するものであってもコミュニケーションの接続を放棄せずに、あくまでもそこに共通理解を見出そうとし続けるというやり方を採用することによって、コミュニケーションの接続を容易で望ましいものにするような相互行為の場がそこに確保されることになるのだと考えられるのである。そして、このときのより大きな社会の秩序に関わる選択とは、具体的レベルの選択に随伴するメタレベルの選択という位置づけになるはずである。
私たちも、コミュニケーションの安定的再生産を確保しようとして、相手の意向に合わせて交渉をまとめようとすることがあるが、それは個々の相互行為を超え出る社会が選択してしまっている秩序のあり方(と当事者が考えているもの)に従属することにしかならない。それは当事者個人の主体的な選択ではないというだけではなく、そのときの相互行為システムの選択にさえなっていない。具体的には、例えば、当事者相互の利害が対立する問題への対処として私たちが採用するふつうのやり方とは、規則や制度のような社会が選択した基準に従って、そのときの利害の対立を調停するような妥当なプランについての合意を形成しようとするというものである。そして、解決が難しい課題に仲間と協働して対処しようとするときに私たちが採用するふつうのやり方とは、そのときのコミュニケーションに先立って存在している「組織」を前提にした集団的意思決定によるものであり、その場合には組織のすべてのメンバーがその決定に従属せざるを得なくなるというものになる。これらのものは、同じ場所に居合わせてコミュニケーションしている人々の主体的な参与による相互行為システムの選択とは、明らかに別のものになる。
その一方で、集団的意思決定を容易にする「組織」を持たず、規則や制度のような社会という個々の相互行為を超え出るまとまりが用意する支えが未発達な、人間以前の社会や人間の原初的な社会では、相互行為システムにおける二重の選択という現象は、ごく一般的なものである。哺乳類の群居的な社会においては、それぞれの個体が一般的な行動傾向として、近くにいる同種個体と活動を同調させようとしながら可能な限りコンフリクトを回避しようとするというやり方を採用することで、そのような状態が繰り返し再生産されるという社会秩序が確保され、それによって同種個体間でのこの種の行為接続が、個別的な出来事として容易に選択できるようになっている。真猿類の群れ社会では、同じ群れのメンバーどうしという関係で活動の同調とコンフリクトの回避を行うことによって、群れというまとまりが維持されてコミュニケーションの安定的再生産を保証する秩序が成立し、その秩序のもとで同じ群れのメンバー間での様々なタイプの行為接続がより容易に選択できるようになっているのである。
チンパンジー属の社会では、同じ集団の他のメンバーと離れて暮らせるようになっているが、離れて暮らしていた個体間で同じ集団のメンバーどうしという関係にあることを確認する手段と機会が用意されていることから、それらを活用することによって同じ集団のメンバーどうしのコミュニケーションの安定的再生産を保証する秩序が成立しているのだと考えられる。さらに、相互行為システムのコミュニケーションが卓越する原初的な人間社会においても、例えば、狩猟採集民ブッシュマンの社会でも、トゥルカナ同様に、徹底的に個人の自律性を尊重し合うというやり方でコミュニケーションの安定的再生産を確保しようとするメタレベルの選択が、不可欠のものとして実行されている。
すでに述べたように、現代日本に生きる私たちにとっては、相互行為システムのコミュニケーションにおいて自らが当事者として、個々の相互行為を超え出るより大きな社会の秩序に関わる選択に直接関与するという経験は、ごく特別なものになってしまっているのだと思える。しかし、以下に示すような相互行為システムのコミュニケーションが重要な機能を果たしている地域社会においては、トゥルカナ社会において私が見出したものと類似の現象が成立しているのではないかと思える。具体的に検討してみよう。
4.島社会における「島で暮らし続ける」という選択
現代日本では、生まれ育った島を出て本土で暮らすということが少しも特別な選択ではなくなってきている。そのような状況では、そこで暮らし続けようとすることは、より積極的な選択として自ら肯定できるものにならなければならなくなっている。そして、「島で暮らし続ける」という選択とは、そのように選択した者にとっては、たんに個人として島から出ないというだけのことではなく、この島の地域社会をこれから先も存続し続けるものにしようとするという覚悟のもとでそうすることになるのだと考えられるのである。それは、「地域社会の消滅」という最悪の事態を起こりうる未来として自覚したうえで、日々の問題対処のための相互行為を実行しながら、同時に、そのときの選択が個々の相互行為を超え出る地域的なコミュニケーションのまとまりの再生産をより容易にするものになっているかを繰り返し反省するようになるということである。
以下では、以前に私が調査した岡山県白石島の事例を手がかりに考察を進めよう。その研究では島社会が抱えるさまざまな問題を取り上げたが、ここでは地域の伝統芸能である「白石踊り」の継承の巡る問題に焦点を当てて検討する。
この白石踊りは、かつては島民のほとんどが踊れた地域の盆踊りであったが、1976年に国の重要無形民俗文化財に指定され、近年は観光の目玉とも目されており、「白石島の誇り」として島民に意識されてもいる。しかし、過疎化による人口減少と、娯楽の多様化による島民の踊り離れもあって、島民のための盆踊りの輪は小規模化し、以前のような盛り上がりは期待すべくもないものになっている。このような状態はかなり以前から誰の目にも明らかなものになっており、伝統行事継承への危機感から、小中学校で子供たちに踊りを教えるという試みが20年以上も続けられている。また、お盆が近づく時期には、観光客向けに踊りの機会を設けたり、チャーター船を仕立てた鑑賞体験ツアーを企画して、一緒に踊りを体験できるようにレクチャーする機会を提供してもいる。
このような伝統行事継承のための活動は、「白石踊会」という島民の中でもとくに熱心な人たちによって担われているが、その活動に対する他の島民の反応は、予想外に懐疑的なものなのである。観光客向けの踊りでは、揃いの衣装で見栄えの良いものが披露されるとともに、踊りの最中に解説が挿入されて中断したりするが、それに対して「これでは誰のための白石踊りかわからない」という不満が述べられる。また、学校教育の場での踊りの指導に対しても、島民にとっての踊りのあるべき姿という観点から、「否応なく習うものになった」という否定的な評価が述べられたりもする。文化財指定という肩書に対してさえ、「文化財に指定されたから伝承しているのではない」という島の人びとの「自分たちの踊り」への誇りが述べられたりもするのである。
しかし、だからといって、外部からの高い評価や経済的効果それ自体を否定的に考えているわけではない。それらは、地元の年中行事として自分たちが大切にしてきたという思いと共に、継承の危機に際してそのあるべき姿を意識することを動機づける重要な要素になっているのである。外部からの要請にも応えながら、その行事のあるべき姿との間に折り合いをつけようと試行錯誤を繰り返してきているのである。このようなやり方で白石島の人びとが実現しようとしていることとは、この行事を今後とも長期間にわたって維持することを可能にするような地域社会の秩序を確保することなのだと考えられる。この場合人びとは、この行事に対する個人的な好みの違いから誰もが同じように積極的に関与するとは限らないということを前提に、それぞれがそれぞれの立場でこの問題に取り組もうとしたときに、それを多くの島民が納得するものへとまとめ上げようとコミュニケーションを接続し続けることが不可欠となるが、それをより容易にするような秩序を確保しようとしているということなのである。
ここで白石島の人びとがしていることは、相互行為システムがする選択という操作それ自体について反省することによって、その前提となる、個々の相互行為を超え出る地域社会の秩序をある望ましいものとして確保しようとすることそのものだと考えられる。
5.おわりに
地域的な広がりの内部におけるコミュニケーションの接続をより容易なものとして確保しようするメタレベルの選択とは、個人の自律的な選択と矛盾しない、共に暮らす仲間との協働による「われわれの選択」によって、すなわち、相互行為システムの主体的な選択によって自らの人生を切り開こうとする生き方にとって、必要不可欠なものになると考えられるのである。ただし、近代に生きる私たちの場合は、仲間との協働による集団的な問題対処において、外部的な権威や制度が指定するものに従属することで仲間との協働が可能になっていることがふつうなのである。その場合には、「相互行為システムの主体的な選択」が問題になることはなく、この種のメタレベルの選択に目が向けられることもない。
ここで注目したようなメタレベルの選択が顕在化することは、相互行為システムのコミュニケーションが当事者の一方が拒否するだけで存在することをやめてしまう状況システムであるという事実に直接由来している。コミュニケーションの地域的まとまりを強化する制度的支えや権威構造を欠いている自然発生的な地域社会では、相互行為システムのコミュニケーションが卓越する一方で、そこでのコミュニケーションの接続が簡単に途絶しないようにしようとして、この種の選択が不可避的に顕在化するのだと考えられる。また、現代日本の島社会をはじめとする消滅可能性が取りざたされる地域社会では、従来からあった組織的・制度的支えの弱体化に伴い、相互行為システムのコミュニケーションの重要性が無視できないものになるのであり、地域的な広がりにおけるこの種のコミュニケーションの接続を確保しようとする選択がより強調された形で顕在化することになるのだと考えられるはずである。
公開・合評会シンポジウム『他者—人類社会の進化』(河合香吏編,京都大学学術出版会,2016)をめぐって
内容:
東京外国語大学アジア・アフリカ言語文化研究所・基幹研究人類学班では,班員の刊行した図書について,内外からコメンテーターをお呼びして合評会(シンポジウム)を開催してきた。今回は,2016年3月に刊行された河合香吏編『他者—人類社会の進化』(京都大学学術出版会)(以下「本書」とする)をとりあげた。帯に記された本書の内容は以下の通りである。
今日「他者」は諸学問の流行テーマである。しかし本書はそれらの議論とは一線を画す。すなわち,一切の思弁を排し,ヒトとサル(そして他の動物)の参与的な観察事例にこだわった厳密な経験科学として,「他者」なるものを析出していく。哲学的な思索の対象としてではなく,個体と個体(集団と集団)の相互行為のプロセスとしての「他者」の中に,人類の社会性の本質を見る。
合評会シンポジウム当日は,開会の挨拶として人類学班の代表である西井凉子(AA研)から人類学班の活動や合評会シンポジウムについての簡単な紹介があり,引き続き,本シンポジウムの企画者である河合香吏(AA研)に司会を移して,以降,河合が進行役を担当した。
はじめに本書の編者である河合が,その概要について簡潔に紹介した。本書はAA研の共同研究課題「人類社会の進化史的基盤研究(3)」の成果論文集として刊行されたものであり,執筆陣の専門は霊長類社会学,生態人類学、社会文化人類学という3学問分野にわたる。本書に収められた各論攷は,ヒトやヒト以外の霊長類における個体間の具体的なインタラクティヴな行為・行動の詳細に着目し,そこに「他者」の出現のありようをみようとするといった,経験科学に立脚した議論が交わされているという特徴がある。それらはまた,「他者とともに生きる」術,すなわち「社会性sociality」を進化の過程で発達させてきた群居性生物としてのヒトを含む霊長類種について,進化的な基盤にまで及んだ議論を展開しているという特徴を併せ持つ。こうした特徴が説明された後,本書の基本的な立ち位置は,緻密で慎重な観察をはじめとする経験論的なアプローチと大胆な抽象的理論化の間を行きつ戻りつしつつ,「他者」の問題に根源的に迫るといったものであることが示された。
続いて,本書の執筆陣から,上記 3分野ごとに,それぞれ内容の紹介がされた。
まず,霊長類学の分野から西江仁徳(京都大学)は,霊長類学では「他者」が問題にされたことはこれまでになかったこと,本書の各章は「普通の(生物学の下位分野としての)霊長類学の論文」とは言い難く,「思弁的霊長類学」などと称されることもあること,それ故に何を書いても新しいものになる一方で,現在の霊長類学の中では評価されにくく,むしろ人類学者など他の分野の読者からの評価を期待するものであるということを主張した。その上で,霊長類学者が他者を論じる意味,すなわちその貢献として,他者の共時的拡張(存在論的拡張)および他者の通時的拡張(進化論的拡張)といった視点を投じることが可能であることを示した。さらに,他者を論じる際の課題として,相互行為を詳細に記述・分析することの徹底や,既存の形式に収まらない表現の工夫=霊長類社会を記述する新たな方法の模索が必要とされること,そして,進化(生物学的な基盤)の視点として「他者的なもの」の進化(起原や変遷)や,人類進化史における「他者的なもの」が与えた影響等を論じる姿勢が今後,求められるであろうこと等が指摘された。
次に,生態人類学の分野から北村光二(岡山大学名誉教授)が,生態人類学における2つのトピックスとして,①人間以前の社会との連続性(ヒトとヒト以外の霊長類をつなぐ「進化」の文脈),および②人間社会における生業ごとのコミュニケーションのありようの違い(人間の生存を支えているものとしての生業を重視する文脈)を挙げ、これらに沿って,生態人類学分野の各論攷を紹介した。いずれの文脈においても,「他者」とは,(本書終章の船曳論文の言うところの)「可能性として第二者になりうる存在」としての他者である。①の文脈においては,哺乳類の群居的な生活形の社会における「社会的促進(活動の枠組みの共有)」と「社会的抑制(敵対的衝突の回避)」という原則が継承されていることが基本となっている相互行為のコミュニケーションの進化について説明され,②の文脈においては,「コミュニケーションの相手として想定される他者」に関わる態度が,狩猟採集,牧畜,農耕といった生業ごとに異なるといった現象があることについて説明された。
最後に,社会文化人類学の分野から船曳建夫(東京大学名誉教授)が,まず,「サル屋(霊長類の研究者)」と「ヒト屋(人間の研究者)」の共同研究の困難さを指摘しつつ,それ故にこうした共同研究は滅多にないのだが,そうした困難が,それぞれのメンバーにとって自省的,反省的になれるという点においてよい機会をもたらす点に触れた。その上で,本書に収められた社会文化人類学分野の6本の論文について個別に紹介しながら,そのすべてに共通するあるまとまった結論が見出せるとした。すなわち,他者はそこにあるものではなく,生み出すもの,生み出されるものであり,そうした他者が生み出される過程に社会性が出現し,それによって社会が生まれるという,そのことである。私という立場から語ると,まず,①私は他個体と共在しながら生きている,②他個体は私と関わりあいができて,その社会関係の形成,社会性の発現の内に,形はさまざまであるが,他者となって現れる,③そこに社会が成立してくるという論理的なプロセスがこの6つの論文によって示されているのだということである。
ここで休憩をとった後,引きつづき,同様に上記3つの学問分野それぞれに与するコメンテーターからコメントをいただいた。
まず,霊長類学の立場からのコメンテーターであるデイビッド・スプレイグ氏(農業・食品産業技術総合研究機構)は,はじめに本書全体に関するコメントとして,大部の本の割には読みやすかったこと,他者論の多様性と奥深さをみせてもらったこと,ひとつの共通理解として「社会交渉の中から他者が生成されていく」という過程があることを挙げ,そして霊長類学の役割はその過程を観察によって記述していくことにあると指摘した。また,本書の読み方として,できるだけ早く読み通すことを勧め,そうすることによってひとつひとつの論文を読んだだけではみえてこない大きな議論の流れ、すなわち一冊の本としての全体像が立体的に把握できると評価した。一方,本書を日本語で読んだことの意義に触れ,日本語の「他者」と英語の’other’の違いから,「他者」は日本語でこそよりクリアに把握できる概念である可能性を示唆した。加えて,まだあまり生物学的ではなかった時代の日本霊長類学について触れ,その頃からの伝統として,ヒトとヒト以外の霊長類を同じ土俵に乗せることがふつうにあったこと,そこでは極めて細かい社会交渉とそこにみえる社会関係の記述に非常にこだわってきたことを指摘し,さらにそこから社会構造の解明へと至る理論構築のありかたについて,本書において再体験できたと感想を述べた。また,このような分野横断的な研究者の集まりは世界に類をみないと強調した。最後に「注文」として,①「者」という言葉(英語にはない)の分析(何故「人」ではなく「者」なのか),②生物学的「個体」論と他者論の関連—個体は他者を前提とするのか,他者のいない個体は可能なのか,等,③自己認識と他者認識の関連,④生物学の哲学について,追究を深めて欲しいとの期待がつけ加えられた。
次に,生態人類学の立場からのコメンテーターとして大石高典氏(東京外国語大学)は,まず,自身のフィールド経験や研究活動経験において,さまざまな契機で「自集団の中の他者」と「他集団の中の他者」の交錯が起きることに接してきたため,本書を自分のしてきたことと関わりのあるものとして親近感を持って読んだと感想を述べた上で,本書に収められた生態人類学分野の論文4本について,ひとつひとつ丁寧にコメントを加え,疑問点や理解できなかった点等について率直な指摘をした。全体については,本書を読めば読むほど「他者」についてわからなくなるという点で「よかった」と感想を述べ,ただし,「他者」といったときに「不可解である」,「パージされる」,あるいは伊谷純一郎のいうところの「許されない」といったニュアンスとの関連が不明瞭であること,使われている進化の概念やイメージが各執筆者間でバラバラであり,この多様性をどのように読んだらよいのか—例えば,事例で扱われている極めて微視的な社会行動と進化的時間との間に飛躍があるように感じられるが、果たしてイメージを膨らませればそれでよいということなのか等,疑問が残るとした。一方で,経験科学の立場から対象に自分を重ね合わせながら研究するというアプローチの可能性が豊かに示されており,相互行為に着目することで種を越えた社会性の研究の展望ができる点にはポジティヴな印象を持ったといい,さらに最近の文化人類学のマルチスピーシーズ人類学における「種」,もの,環境を扱う潮流とシンクロナイズしているのかもしれないとの感想を述べた。
最後に,佐久間寛氏(AA研)は,「社会文化人類学的苦悩あるいは可能性としての表象/想像力—他者研の他者からのコメント」と題してコメントした。まず,共同研究課題「人類社会の進化史的基盤研究」が進化しつつも退化した部分(議論から抜け落ちていった部分)もあるとし,これを表象/想像力という問題であるとした。とくに本書の第3部(主として社会文化人類学の論文を集めた部)では,表象能力や想像力について語られてもよいはずだが,これらは後背に退いており,むしろ存在論的な議論が主題として展開されているとみる。そして,これを表象論に対する近年の存在論的批判,すなわち,人類学の「静かな革命」,「存在論的転回」等で語られ,文化人類学における大きな潮流となっているPh.デスコラやヴィヴェイロス・デ・カストロ,ラトゥールらの議論と(本書ではこれらを直接引用している論文は決して多くはないものの),軌を一にしているように感じられると指摘した。さらに,他者をめぐる代理表象(認識論的暴力)の問題には,本書では人類学者よりも霊長類学者のほうが,よりビビッドで敏感に反応していることに言及した。そして,主体性の問題にも深く関わるこうした他者に対する表象の問題にも本書でもっと応答して欲しかったと述べた。次に挙げられた論点は「人類学者という他者」の問題であり,人類学者がフィールドに行って,どのような他者性をどのように帯びてゆくのかという問題の提議であった。最後に,他者を考えるに当たって人類と霊長類の境界(あるいは断絶)という問題に対し,とりわけ社会文化人類学分野の執筆者が向き合っていない点に物足りなさを感じたと評した。
以上,本書の内容紹介とコメントとのやりとりを踏まえ,この後1時間ほどを費やして,フロアの参加者をも含めて活発な討論が展開された。
今日「他者」は諸学問の流行テーマである。しかし本書はそれらの議論とは一線を画す。すなわち,一切の思弁を排し,ヒトとサル(そして他の動物)の参与的な観察事例にこだわった厳密な経験科学として,「他者」なるものを析出していく。哲学的な思索の対象としてではなく,個体と個体(集団と集団)の相互行為のプロセスとしての「他者」の中に,人類の社会性の本質を見る。
合評会シンポジウム当日は,開会の挨拶として人類学班の代表である西井凉子(AA研)から人類学班の活動や合評会シンポジウムについての簡単な紹介があり,引き続き,本シンポジウムの企画者である河合香吏(AA研)に司会を移して,以降,河合が進行役を担当した。
はじめに本書の編者である河合が,その概要について簡潔に紹介した。本書はAA研の共同研究課題「人類社会の進化史的基盤研究(3)」の成果論文集として刊行されたものであり,執筆陣の専門は霊長類社会学,生態人類学、社会文化人類学という3学問分野にわたる。本書に収められた各論攷は,ヒトやヒト以外の霊長類における個体間の具体的なインタラクティヴな行為・行動の詳細に着目し,そこに「他者」の出現のありようをみようとするといった,経験科学に立脚した議論が交わされているという特徴がある。それらはまた,「他者とともに生きる」術,すなわち「社会性sociality」を進化の過程で発達させてきた群居性生物としてのヒトを含む霊長類種について,進化的な基盤にまで及んだ議論を展開しているという特徴を併せ持つ。こうした特徴が説明された後,本書の基本的な立ち位置は,緻密で慎重な観察をはじめとする経験論的なアプローチと大胆な抽象的理論化の間を行きつ戻りつしつつ,「他者」の問題に根源的に迫るといったものであることが示された。
続いて,本書の執筆陣から,上記 3分野ごとに,それぞれ内容の紹介がされた。
まず,霊長類学の分野から西江仁徳(京都大学)は,霊長類学では「他者」が問題にされたことはこれまでになかったこと,本書の各章は「普通の(生物学の下位分野としての)霊長類学の論文」とは言い難く,「思弁的霊長類学」などと称されることもあること,それ故に何を書いても新しいものになる一方で,現在の霊長類学の中では評価されにくく,むしろ人類学者など他の分野の読者からの評価を期待するものであるということを主張した。その上で,霊長類学者が他者を論じる意味,すなわちその貢献として,他者の共時的拡張(存在論的拡張)および他者の通時的拡張(進化論的拡張)といった視点を投じることが可能であることを示した。さらに,他者を論じる際の課題として,相互行為を詳細に記述・分析することの徹底や,既存の形式に収まらない表現の工夫=霊長類社会を記述する新たな方法の模索が必要とされること,そして,進化(生物学的な基盤)の視点として「他者的なもの」の進化(起原や変遷)や,人類進化史における「他者的なもの」が与えた影響等を論じる姿勢が今後,求められるであろうこと等が指摘された。
次に,生態人類学の分野から北村光二(岡山大学名誉教授)が,生態人類学における2つのトピックスとして,①人間以前の社会との連続性(ヒトとヒト以外の霊長類をつなぐ「進化」の文脈),および②人間社会における生業ごとのコミュニケーションのありようの違い(人間の生存を支えているものとしての生業を重視する文脈)を挙げ、これらに沿って,生態人類学分野の各論攷を紹介した。いずれの文脈においても,「他者」とは,(本書終章の船曳論文の言うところの)「可能性として第二者になりうる存在」としての他者である。①の文脈においては,哺乳類の群居的な生活形の社会における「社会的促進(活動の枠組みの共有)」と「社会的抑制(敵対的衝突の回避)」という原則が継承されていることが基本となっている相互行為のコミュニケーションの進化について説明され,②の文脈においては,「コミュニケーションの相手として想定される他者」に関わる態度が,狩猟採集,牧畜,農耕といった生業ごとに異なるといった現象があることについて説明された。
最後に,社会文化人類学の分野から船曳建夫(東京大学名誉教授)が,まず,「サル屋(霊長類の研究者)」と「ヒト屋(人間の研究者)」の共同研究の困難さを指摘しつつ,それ故にこうした共同研究は滅多にないのだが,そうした困難が,それぞれのメンバーにとって自省的,反省的になれるという点においてよい機会をもたらす点に触れた。その上で,本書に収められた社会文化人類学分野の6本の論文について個別に紹介しながら,そのすべてに共通するあるまとまった結論が見出せるとした。すなわち,他者はそこにあるものではなく,生み出すもの,生み出されるものであり,そうした他者が生み出される過程に社会性が出現し,それによって社会が生まれるという,そのことである。私という立場から語ると,まず,①私は他個体と共在しながら生きている,②他個体は私と関わりあいができて,その社会関係の形成,社会性の発現の内に,形はさまざまであるが,他者となって現れる,③そこに社会が成立してくるという論理的なプロセスがこの6つの論文によって示されているのだということである。
ここで休憩をとった後,引きつづき,同様に上記3つの学問分野それぞれに与するコメンテーターからコメントをいただいた。
まず,霊長類学の立場からのコメンテーターであるデイビッド・スプレイグ氏(農業・食品産業技術総合研究機構)は,はじめに本書全体に関するコメントとして,大部の本の割には読みやすかったこと,他者論の多様性と奥深さをみせてもらったこと,ひとつの共通理解として「社会交渉の中から他者が生成されていく」という過程があることを挙げ,そして霊長類学の役割はその過程を観察によって記述していくことにあると指摘した。また,本書の読み方として,できるだけ早く読み通すことを勧め,そうすることによってひとつひとつの論文を読んだだけではみえてこない大きな議論の流れ、すなわち一冊の本としての全体像が立体的に把握できると評価した。一方,本書を日本語で読んだことの意義に触れ,日本語の「他者」と英語の’other’の違いから,「他者」は日本語でこそよりクリアに把握できる概念である可能性を示唆した。加えて,まだあまり生物学的ではなかった時代の日本霊長類学について触れ,その頃からの伝統として,ヒトとヒト以外の霊長類を同じ土俵に乗せることがふつうにあったこと,そこでは極めて細かい社会交渉とそこにみえる社会関係の記述に非常にこだわってきたことを指摘し,さらにそこから社会構造の解明へと至る理論構築のありかたについて,本書において再体験できたと感想を述べた。また,このような分野横断的な研究者の集まりは世界に類をみないと強調した。最後に「注文」として,①「者」という言葉(英語にはない)の分析(何故「人」ではなく「者」なのか),②生物学的「個体」論と他者論の関連—個体は他者を前提とするのか,他者のいない個体は可能なのか,等,③自己認識と他者認識の関連,④生物学の哲学について,追究を深めて欲しいとの期待がつけ加えられた。
次に,生態人類学の立場からのコメンテーターとして大石高典氏(東京外国語大学)は,まず,自身のフィールド経験や研究活動経験において,さまざまな契機で「自集団の中の他者」と「他集団の中の他者」の交錯が起きることに接してきたため,本書を自分のしてきたことと関わりのあるものとして親近感を持って読んだと感想を述べた上で,本書に収められた生態人類学分野の論文4本について,ひとつひとつ丁寧にコメントを加え,疑問点や理解できなかった点等について率直な指摘をした。全体については,本書を読めば読むほど「他者」についてわからなくなるという点で「よかった」と感想を述べ,ただし,「他者」といったときに「不可解である」,「パージされる」,あるいは伊谷純一郎のいうところの「許されない」といったニュアンスとの関連が不明瞭であること,使われている進化の概念やイメージが各執筆者間でバラバラであり,この多様性をどのように読んだらよいのか—例えば,事例で扱われている極めて微視的な社会行動と進化的時間との間に飛躍があるように感じられるが、果たしてイメージを膨らませればそれでよいということなのか等,疑問が残るとした。一方で,経験科学の立場から対象に自分を重ね合わせながら研究するというアプローチの可能性が豊かに示されており,相互行為に着目することで種を越えた社会性の研究の展望ができる点にはポジティヴな印象を持ったといい,さらに最近の文化人類学のマルチスピーシーズ人類学における「種」,もの,環境を扱う潮流とシンクロナイズしているのかもしれないとの感想を述べた。
最後に,佐久間寛氏(AA研)は,「社会文化人類学的苦悩あるいは可能性としての表象/想像力—他者研の他者からのコメント」と題してコメントした。まず,共同研究課題「人類社会の進化史的基盤研究」が進化しつつも退化した部分(議論から抜け落ちていった部分)もあるとし,これを表象/想像力という問題であるとした。とくに本書の第3部(主として社会文化人類学の論文を集めた部)では,表象能力や想像力について語られてもよいはずだが,これらは後背に退いており,むしろ存在論的な議論が主題として展開されているとみる。そして,これを表象論に対する近年の存在論的批判,すなわち,人類学の「静かな革命」,「存在論的転回」等で語られ,文化人類学における大きな潮流となっているPh.デスコラやヴィヴェイロス・デ・カストロ,ラトゥールらの議論と(本書ではこれらを直接引用している論文は決して多くはないものの),軌を一にしているように感じられると指摘した。さらに,他者をめぐる代理表象(認識論的暴力)の問題には,本書では人類学者よりも霊長類学者のほうが,よりビビッドで敏感に反応していることに言及した。そして,主体性の問題にも深く関わるこうした他者に対する表象の問題にも本書でもっと応答して欲しかったと述べた。次に挙げられた論点は「人類学者という他者」の問題であり,人類学者がフィールドに行って,どのような他者性をどのように帯びてゆくのかという問題の提議であった。最後に,他者を考えるに当たって人類と霊長類の境界(あるいは断絶)という問題に対し,とりわけ社会文化人類学分野の執筆者が向き合っていない点に物足りなさを感じたと評した。
以上,本書の内容紹介とコメントとのやりとりを踏まえ,この後1時間ほどを費やして,フロアの参加者をも含めて活発な討論が展開された。
(文責:河合香吏)
第5回研究会での発表内容
1.「変動する生息環境とチンパンジーの生存」(伊藤詞子)
本発表では、東アフリカ西端に位置するタンザニア連合共和国マハレ山塊国立公園の、あるチンパンジー(Pan troglodytes)集団の生を支える、生息環境について、気候、植物フェノロジー、植生について、特に中・長期的変動に着目して、発表をおこなった(詳しくは以下の通り)。
1. 気候(Itoh 2015)
マハレの気候は年毎にも、10年単位でも、変動が大きいことで知られる。こうした周期的変動と長期的変動を区別するためには、30年以上の資料が必要といわれる。1983年から2013年までの、マハレに滞在していた研究者等が、毎朝記録し、蓄積してきた気象データから、以下の長期的変動が推測されている。
1-1 雨量(N=11,142日):マハレはタンザニアでも有数の多雨地帯であり、31年間の平均年間降雨量は1,751ミリメートルである。各日の平均からの差(365日間の移動平均)から31年間の傾向を確認したところ、7.7ミリメートル/年の減少傾向が見られた。さらに、31年分のデータを、前半と後半に二分して比較したところ、雨期の前半に雨が増え、後半に減少する傾向が見られた。
1-2 気温:各日の平均からの差(31日間の移動平均)から31年間の傾向を確認したところ、最低気温(N=10,476)、最高気温(N=10,337)ともに温暖化傾向が推測された。すなわち、前者は変異幅が小さく摂氏±1度を超えることは滅多になく、その一方で、最高気温の変異幅は摂氏8度(+5℃〜-3℃)と大きかった。
2. 植物フェノロジー(季節動態)
マハレでは現地調査助手等を中心に、1997年からライン・トランゼクト上にある、木本の樹木とツル個体の、開花・結実・展葉について毎月一回毎木調査をおこなっている。これらの資料のうち、チンパンジーの重要な食物である17種(Itoh and Nakamura 2015a)について、1997年から2013年までのデータを解析した(Itoh and Muramatsu 2015)。その結果、2005年前後を境に、特に年単位周期が認められる種において、結実動態が変化してきている可能性が示唆された。また、17種中年によらず最もよく利用されている7種のほとんどは、結実の年単位での周期性が明瞭な種がほとんどであった。
3. 植生(Itoh and Nakamura 2015b)
マハレの植生は、もともとこの一帯に住んでいた地元住民の居住が禁止されて以降、徐々に遷移が進んでいる。1990年代終わりにおこなったトランゼクト(上記の植物フェノロジーも同じトランゼクトでおこなっている)において、2010年代初めに再度植生調査をおこなわれている。トランゼクトは低地林帯にあり、トランゼクトの中央から両側各5メートル内にある、胸高直径10センチメートル以上の樹木と、5センチメートル以上の木本ツル植物個体すべてを対象にしている。トランゼクトの長さは、初めの調査で8.38キロメートルであった。しかし、10年余りの月日の流れの中で、樹木が転倒したり、大木の樹幹が落ちたり、あるいは川の流れが変わるなどの影響によって、道そのものがあちこちでそうした障害を迂回していたため、二度目の調査で完全にトランゼクトを一致させることはできなかった。樹木の相対的位置関係から、完全に一致していると推測された区画(1区画=10メートル長×10メートル幅)内の木本植物の密度、樹木の胸高直径と基底面積などを比較した(総面積6.63ヘクタール)。その結果、以下の点が明らかになっている。まず、樹木の基底面積や胸高直径分布などの指標から、森林は大きくなっていると推測された。また遷移の過程で優先すると推測されていたバンレイシ科のXylopiaには、増加傾向が見られた。しかし、当初予測されていた遷移過程とは異なるプロセスとして、パイオニア種であるニレ科のTrema orientalis (L.) Blumeが依然として多いこと、また、木本のツル植物の増加などがあげられる。ツルが森林に占める割合は26.4%から32.1%に増加したが、ツルの中でも、特にチンパンジーの最重要果実食物であるキョウチクトウ科のSaba comorensis (Bojer ex A.DC.) Pichonの増加傾向が顕著であった。また、1980年代以降のグローバルな熱帯林の変動傾向として、樹木のターンオーバー率が高くなってきていることが報告されている。マハレの樹木のターンオーバー率は、そうした世界中の熱帯林で認められている値に匹敵する(旧大陸1.81±0.19%, 新大陸1.80±0.15%, マハレ1.61%)。
マハレの気候は数年〜十年単位での変動が大きいことでも知られており、植物フェノロジーや植生の調査には、さらなる資料の蓄積が必須である。一方で、植物にかかわる資料によって現在認められているいくつかの傾向は、世界中の熱帯林でも観察されており、地球規模の気候変動の影響が示唆されている。こうした変動は、チンパンジーの生息環境としては好ましからざる傾向も推測される。しかし、長期にわたるこうした漸進的変化に対し、チンパンジーが即座に行動変化をおこしているという明確な証拠はない。今後、チンパンジーの「移動」という観点から、明らかになっている環境変動の再検討をおこなう。
引用文献
1. 気候(Itoh 2015)
マハレの気候は年毎にも、10年単位でも、変動が大きいことで知られる。こうした周期的変動と長期的変動を区別するためには、30年以上の資料が必要といわれる。1983年から2013年までの、マハレに滞在していた研究者等が、毎朝記録し、蓄積してきた気象データから、以下の長期的変動が推測されている。
1-1 雨量(N=11,142日):マハレはタンザニアでも有数の多雨地帯であり、31年間の平均年間降雨量は1,751ミリメートルである。各日の平均からの差(365日間の移動平均)から31年間の傾向を確認したところ、7.7ミリメートル/年の減少傾向が見られた。さらに、31年分のデータを、前半と後半に二分して比較したところ、雨期の前半に雨が増え、後半に減少する傾向が見られた。
1-2 気温:各日の平均からの差(31日間の移動平均)から31年間の傾向を確認したところ、最低気温(N=10,476)、最高気温(N=10,337)ともに温暖化傾向が推測された。すなわち、前者は変異幅が小さく摂氏±1度を超えることは滅多になく、その一方で、最高気温の変異幅は摂氏8度(+5℃〜-3℃)と大きかった。
2. 植物フェノロジー(季節動態)
マハレでは現地調査助手等を中心に、1997年からライン・トランゼクト上にある、木本の樹木とツル個体の、開花・結実・展葉について毎月一回毎木調査をおこなっている。これらの資料のうち、チンパンジーの重要な食物である17種(Itoh and Nakamura 2015a)について、1997年から2013年までのデータを解析した(Itoh and Muramatsu 2015)。その結果、2005年前後を境に、特に年単位周期が認められる種において、結実動態が変化してきている可能性が示唆された。また、17種中年によらず最もよく利用されている7種のほとんどは、結実の年単位での周期性が明瞭な種がほとんどであった。
3. 植生(Itoh and Nakamura 2015b)
マハレの植生は、もともとこの一帯に住んでいた地元住民の居住が禁止されて以降、徐々に遷移が進んでいる。1990年代終わりにおこなったトランゼクト(上記の植物フェノロジーも同じトランゼクトでおこなっている)において、2010年代初めに再度植生調査をおこなわれている。トランゼクトは低地林帯にあり、トランゼクトの中央から両側各5メートル内にある、胸高直径10センチメートル以上の樹木と、5センチメートル以上の木本ツル植物個体すべてを対象にしている。トランゼクトの長さは、初めの調査で8.38キロメートルであった。しかし、10年余りの月日の流れの中で、樹木が転倒したり、大木の樹幹が落ちたり、あるいは川の流れが変わるなどの影響によって、道そのものがあちこちでそうした障害を迂回していたため、二度目の調査で完全にトランゼクトを一致させることはできなかった。樹木の相対的位置関係から、完全に一致していると推測された区画(1区画=10メートル長×10メートル幅)内の木本植物の密度、樹木の胸高直径と基底面積などを比較した(総面積6.63ヘクタール)。その結果、以下の点が明らかになっている。まず、樹木の基底面積や胸高直径分布などの指標から、森林は大きくなっていると推測された。また遷移の過程で優先すると推測されていたバンレイシ科のXylopiaには、増加傾向が見られた。しかし、当初予測されていた遷移過程とは異なるプロセスとして、パイオニア種であるニレ科のTrema orientalis (L.) Blumeが依然として多いこと、また、木本のツル植物の増加などがあげられる。ツルが森林に占める割合は26.4%から32.1%に増加したが、ツルの中でも、特にチンパンジーの最重要果実食物であるキョウチクトウ科のSaba comorensis (Bojer ex A.DC.) Pichonの増加傾向が顕著であった。また、1980年代以降のグローバルな熱帯林の変動傾向として、樹木のターンオーバー率が高くなってきていることが報告されている。マハレの樹木のターンオーバー率は、そうした世界中の熱帯林で認められている値に匹敵する(旧大陸1.81±0.19%, 新大陸1.80±0.15%, マハレ1.61%)。
マハレの気候は数年〜十年単位での変動が大きいことでも知られており、植物フェノロジーや植生の調査には、さらなる資料の蓄積が必須である。一方で、植物にかかわる資料によって現在認められているいくつかの傾向は、世界中の熱帯林でも観察されており、地球規模の気候変動の影響が示唆されている。こうした変動は、チンパンジーの生息環境としては好ましからざる傾向も推測される。しかし、長期にわたるこうした漸進的変化に対し、チンパンジーが即座に行動変化をおこしているという明確な証拠はない。今後、チンパンジーの「移動」という観点から、明らかになっている環境変動の再検討をおこなう。
引用文献
- Itoh Noriko, 2015. Climate and Climatological Trends in the Kasoje Forest. In: Nakamura Michio, Hosaka Kazuhiko, Itoh Noriko, Zamma Koichiro (Eds), Mahale Chimpanzees: 50 Years of Research Cambridge University Press: Cambridge. pp: 143–149
- Itoh Noriko, Muramatsu Daisuke, 2015. Patterns and Trends in Fruiting Phenology: Some Important Implications for Chimpanzee Diet. In: Nakamura Michio, Hosaka Kazuhiko, Itoh Noriko, Zamma Koichiro (Eds), Mahale Chimpanzees: 50 Years of Research Cambridge University Press: Cambridge. pp: 174–194
- Itoh Noriko, Nakamura Michio, 2015a. Diet and Feeding Behavior. In: Nakamura Michio, Hosaka Kazuhiko, Itoh Noriko, Zamma Koichiro (Eds), Mahale Chimpanzees: 50 Years of Research Cambridge University Press: Cambridge. pp: 227–245
- Itoh Noriko, Nakamura Michio, 2015b. Mahale Flora: Its Historical Background and Long-Term Changes. In: Nakamura Michio, Hosaka Kazuhiko, Itoh Noriko, Zamma Koichiro (Eds), Mahale Chimpanzees: 50 Years of Research Cambridge University Press: Cambridge. pp: 150–173
2.極限のなかでリスクから目をそらし続けること(曽我亨)
本発表では、災害(disaster)に関する先行研究を手がかりにしながら、極限(extremity)について考えた。災害の人類学的研究では、破壊素因(ハザード)と人間集団のむすびつきによって災害がおきるとしている。人間集団には様々な脆弱性が内在されており、人間集団の場所・社会基盤・社会政治組織・生産分配体制・イデオロギーなどによって災害の規模や様相が変わることが指摘されている。災害の人類学においては、これらの脆弱性を調べること、また災害が起きた後の社会・文化変容について調べることの重要性が指摘されている。一方、災害研究は、戦争などのように損害を人間が意図的に引き起こすことは研究の対象外であると述べている。
これらを手がかりにして、極限について考えた。まず極限について考える場合は、戦争など、意図的な損害についても対象となり得る。また災害が、破壊素因(ハザード)と人間集団との結びつきによって生じるのに対し、極限の場合は破壊素因ばかりでなく、逆に社会や文化によっても生じうる。そこで本発表では、極限を「自然または人工の環境の変化、あるいは社会状態の変化によって、人々が、これまで取ってきた環境あるいは社会状態への働きかけを持続するだけでは、自らの生存が危機にさらされることを認識し、新たな対応を(その人々が取り得る選択肢の幅の中において)取らなければならない状態」と考えることにした。
本発表がとりあげたのは東アフリカの牧畜民ガブラである。ガブラの人々が直面する極限には、自然の環境変化(旱魃、大雨、疫病)に加え、社会状態の変化(近隣民族との関係悪化、文化的に忌避される事故の発生)によるものが存在する。
自然の環境変化による極限状態としては、1997年のエルニーニョによって生じた大雨と致死性の高いリフトバレー熱の流行、数年に一度おきるマラリアの大流行、また歴史的には1950年代までたびたび流行した天然痘などがある。また19世紀末には、東アフリカ全体を襲った牛疫から逃れていったという記録もある。最近では2010年から12年にかけて東アフリカを大旱魃が襲った。こうした時、人々は旱魃から「逃げ」、よりよい牧草地を探して移動した。自然の環境変化(悪性の)に対しては、逃げるというのが基本的な牧畜民の態度である。
一方、飼育する家畜によっては、自然の変化のスピードについて行けないこともある。たとえば、ラクダを中心に飼育する牧畜民は、旱魃による地域の劣化よりも早く移動することが可能であるが、ウシを中心に飼育する牧畜民は、道中、ウシに給水しない限り、遠くへ移動することはできない。そこでウシ牧畜民は、旱魃から逃げるよりも、むしろ井戸の管理を徹底することで、旱魃をその場でやり過ごそうとする。このように生業経済の差が、極限への対応の違いとして現れることを指摘した。
つぎに社会状態の変化であるが、発表では近隣民族との関係悪化に注目した。ガブラの人々は、20世紀に入ってから今日までに5度の民族紛争を経験し、そのたびに難民・避難民生活を送った。そして紛争が沈静化すると、難民・避難民は故郷に戻り、生活を再建してきた。一般的に、難民が故地に戻り、生活を再建することはとても難しい。ガブラがいとも簡単に再建をなしとげた背景には、家族の一部を故地に残し、家畜の管理を続けたことがあげられる。つまり、敵の襲撃がおきると、ガブラは家族集団を分け、老人などについては生命の安全を確保するため遠くへと逃がし、若者については家畜の世話を続けるため、危険な故地で暮らし続けるのである。このように危険な地域に暮らし続けることを、どのように理解すれば良いのだろうか。
2005年7月、グジ・オロモはボラナ県北部をふくむ地域に新しい県の創出を企てた。しかしこの企ては、ボラナ県のマジョリティであるボラナ・オロモの不興をかうことになった。グジはボラナ県北部への移住をすすめる一方、この地域に住むガブラやボラナを追い出すため、武力攻撃を仕掛けた。多くのガブラやボラナが南方へと逃げ出していった。その一方で、このときも少なからぬ数のガブラがとどまり、家畜の管理を続けていた。
報告者は、2005年8月、ボラナ県北部にとどまった住民数名を、エチオピアとケニアの国境の町モヤレに呼び出し、現地で何が起きているのかインタビュー調査した。そのとき、ある男性は「グジはガブラ全員を滅ぼすといった。しかし、そんなことは絶対にさせない。私たちは戦うつもりだ。もし一人でもガブラが生き残れば、ガブラの勝ちだ」と述べた。ひりひりするような緊張感のなかで彼らが暮らしていることが垣間見られた。
さて、このような発言を、どのように理解したら良いのだろうか。本論ではロバート・ペインのリスク否定の考え方をとりあげ、危険な地域にとどまり続けるガブラの行動について考えた。リスク否定とは、あまりに高いリスクが満ちている状況において、そのリスクを認知的に抑圧することを指す。リスク否定をする人々は、危険があるにもかかわらず、企てたことをあくまで通し続ける。そして熱狂的行動や殉死のように、危険それ自体が価値を生み出すものとして信奉されることもある。
ガブラの行動について検討すると、近隣民族との関係において圧倒的な劣勢におかれながらも、彼らはそこにとどまり続けていた。そして、先に紹介したように、「最後の一人が生き延びれば自分たちの勝利である」というような滅びの美学に通じる発言がみられた。これはリスク否定に登場した殉死の考え方と共通している。また、牧畜民は、家畜を集落から遠く離れた牧草地につれていくことが一般的であるが、こうした家畜キャンプは敵対する民族との境界地域に設置されることも多い。以前、私はガブラの男性から「人間は家畜のために生きるのだ」と聞かされたことがあるが、これもリスク否定の一例となろう。
以上のことから、リスク否定の諸相については引き続き検討する必要があるが、極限状態に身をおき続けようとするガブラの生き方を理解するための補助線となりうることが示唆された。
【参考文献】
これらを手がかりにして、極限について考えた。まず極限について考える場合は、戦争など、意図的な損害についても対象となり得る。また災害が、破壊素因(ハザード)と人間集団との結びつきによって生じるのに対し、極限の場合は破壊素因ばかりでなく、逆に社会や文化によっても生じうる。そこで本発表では、極限を「自然または人工の環境の変化、あるいは社会状態の変化によって、人々が、これまで取ってきた環境あるいは社会状態への働きかけを持続するだけでは、自らの生存が危機にさらされることを認識し、新たな対応を(その人々が取り得る選択肢の幅の中において)取らなければならない状態」と考えることにした。
本発表がとりあげたのは東アフリカの牧畜民ガブラである。ガブラの人々が直面する極限には、自然の環境変化(旱魃、大雨、疫病)に加え、社会状態の変化(近隣民族との関係悪化、文化的に忌避される事故の発生)によるものが存在する。
自然の環境変化による極限状態としては、1997年のエルニーニョによって生じた大雨と致死性の高いリフトバレー熱の流行、数年に一度おきるマラリアの大流行、また歴史的には1950年代までたびたび流行した天然痘などがある。また19世紀末には、東アフリカ全体を襲った牛疫から逃れていったという記録もある。最近では2010年から12年にかけて東アフリカを大旱魃が襲った。こうした時、人々は旱魃から「逃げ」、よりよい牧草地を探して移動した。自然の環境変化(悪性の)に対しては、逃げるというのが基本的な牧畜民の態度である。
一方、飼育する家畜によっては、自然の変化のスピードについて行けないこともある。たとえば、ラクダを中心に飼育する牧畜民は、旱魃による地域の劣化よりも早く移動することが可能であるが、ウシを中心に飼育する牧畜民は、道中、ウシに給水しない限り、遠くへ移動することはできない。そこでウシ牧畜民は、旱魃から逃げるよりも、むしろ井戸の管理を徹底することで、旱魃をその場でやり過ごそうとする。このように生業経済の差が、極限への対応の違いとして現れることを指摘した。
つぎに社会状態の変化であるが、発表では近隣民族との関係悪化に注目した。ガブラの人々は、20世紀に入ってから今日までに5度の民族紛争を経験し、そのたびに難民・避難民生活を送った。そして紛争が沈静化すると、難民・避難民は故郷に戻り、生活を再建してきた。一般的に、難民が故地に戻り、生活を再建することはとても難しい。ガブラがいとも簡単に再建をなしとげた背景には、家族の一部を故地に残し、家畜の管理を続けたことがあげられる。つまり、敵の襲撃がおきると、ガブラは家族集団を分け、老人などについては生命の安全を確保するため遠くへと逃がし、若者については家畜の世話を続けるため、危険な故地で暮らし続けるのである。このように危険な地域に暮らし続けることを、どのように理解すれば良いのだろうか。
2005年7月、グジ・オロモはボラナ県北部をふくむ地域に新しい県の創出を企てた。しかしこの企ては、ボラナ県のマジョリティであるボラナ・オロモの不興をかうことになった。グジはボラナ県北部への移住をすすめる一方、この地域に住むガブラやボラナを追い出すため、武力攻撃を仕掛けた。多くのガブラやボラナが南方へと逃げ出していった。その一方で、このときも少なからぬ数のガブラがとどまり、家畜の管理を続けていた。
報告者は、2005年8月、ボラナ県北部にとどまった住民数名を、エチオピアとケニアの国境の町モヤレに呼び出し、現地で何が起きているのかインタビュー調査した。そのとき、ある男性は「グジはガブラ全員を滅ぼすといった。しかし、そんなことは絶対にさせない。私たちは戦うつもりだ。もし一人でもガブラが生き残れば、ガブラの勝ちだ」と述べた。ひりひりするような緊張感のなかで彼らが暮らしていることが垣間見られた。
さて、このような発言を、どのように理解したら良いのだろうか。本論ではロバート・ペインのリスク否定の考え方をとりあげ、危険な地域にとどまり続けるガブラの行動について考えた。リスク否定とは、あまりに高いリスクが満ちている状況において、そのリスクを認知的に抑圧することを指す。リスク否定をする人々は、危険があるにもかかわらず、企てたことをあくまで通し続ける。そして熱狂的行動や殉死のように、危険それ自体が価値を生み出すものとして信奉されることもある。
ガブラの行動について検討すると、近隣民族との関係において圧倒的な劣勢におかれながらも、彼らはそこにとどまり続けていた。そして、先に紹介したように、「最後の一人が生き延びれば自分たちの勝利である」というような滅びの美学に通じる発言がみられた。これはリスク否定に登場した殉死の考え方と共通している。また、牧畜民は、家畜を集落から遠く離れた牧草地につれていくことが一般的であるが、こうした家畜キャンプは敵対する民族との境界地域に設置されることも多い。以前、私はガブラの男性から「人間は家畜のために生きるのだ」と聞かされたことがあるが、これもリスク否定の一例となろう。
以上のことから、リスク否定の諸相については引き続き検討する必要があるが、極限状態に身をおき続けようとするガブラの生き方を理解するための補助線となりうることが示唆された。
【参考文献】
- ロバート・ペイン2006「危険とリスク否定論」スザンナ・ホフマン, アンソニー・オリヴァー=スミス共編『災害の人類学』明石書店 pp.77-103
3. 「チンパンジーの孤児の生存をめぐって—「母親の不在」は極限的な社会環境か?」(中村美知夫)
ヒトにおいて特別と考えられているいくつかの生活史上の特徴について、野生チンパンジー社会における孤児の存在(母親の不在)から再考する。そのような特徴として頻繁に取り上げられるものとして、早い離乳や幼少期(childhood)・ワカモノ期(adolescence)・祖母期(grandmotherhood)の存在などがある。
ヒトの授乳終了は、非産業社会の民族誌データなどから概ね2.5歳と言われており、一方、野生チンパンジーでは、平均5.2歳まで乳首接触が続く。第一大臼歯の萌出がヒトで6歳頃、チンパンジーで3歳頃であることを考えると、ヒトはかなり早い段階で離乳することになる。
これと連動したヒト特有の発達段階として「幼少期」が考えられている。これは、アカンボウ期とコドモ期の間に位置し、離乳と乳歯列の完成から6.9歳までとされている。幼少期には乳歯と小さな消化器系のため、噛むのが簡単で栄養が密な食物を食べる必要がある。自力では食物獲得ができないため、集団の年長メンバーが食物を集めて与えるということになる。すなわち、幼少期のヒトは自力で生きられず、親をはじめとした年長メンバーに栄養獲得を依存する必要がある。これを裏返せば、類人猿では、離乳さえしてしまえば自力で食物を得て生存できるということになる。
形質人類学では、「ワカモノ期」もまたヒトに特有であるとされる。その場合、ワカモノ期は、急激な成長スパートに特徴づけられる。チンパンジーではそうしたスパートが欠けているために、ヒトに特有であるとされているのだが、霊長類学者の多くは、ヒト以外の霊長類にもワカモノ期があると考えている。その場合は、性成熟から完全な体サイズになるまでといった定義になるが、以下ではワカモノ期についての検討は割愛する。
ヒトの女性は、閉経後(繁殖終了後)も長く生存する。この閉経後の期間が祖母期である。チンパンジーの雌の多くは死ぬまで出産を続けるため、これもまたヒトに特有の生活史段階ということになっている。祖母期の女性は、自分自身では繁殖をせず、自分の子の育児を手伝うことで、包括適応度を上げていると考えられている。幼少期の存在とも密接に関連し、母親が授乳を早く切り上げて、次の子の妊娠・出産にエネルギーを投資できるのも、祖母のような母親以外の養育者が協力するからだとされる。
こうした生活史上の特徴は、その他のさまざまなヒトの特徴と関連づけられて議論される。たとえば、脳が大型化し始めると、母乳だけでは成長に必要なタンパク質が不足する。このため、栄養を肉で補填する必要が生じ、早い段階での離乳が有利になるとする考え方がある。これは、肉食への移行とも関連する。肉の獲得については、かつては狩猟の重要性が強調されていたが、現在ではスキャベンジング(屍肉食)が重要であったと考える研究者が多い。スキャベンジングには、肉食獣が完全に去った後に、残った骨から骨髄などを利用するパッシブ・スキャベンジングと、より積極的に肉食獣を追い払って肉を獲得するパワー・スキャベンジングが考えられている。後者の場合、肉食獣と対峙するため、危険を伴い、若いオトナの死亡率が高くなると予測される。そうすると、子は早い段階で離乳しておいたほうが有利であることになる。
「超−協力(hyper-cooperation)」もまた、ヒトに独特な特徴であると考えられている。複数の霊長類種を対象にした実験で、アロケア(allocare:母以外による子へのケア)の存在と協力行動に連関があるとする結果がある。こうした結果をもとに、早い離乳と幼少期の存在によって協同育児(当然アロケアを伴う)が適応的になり、「超−協力」が進化したと考えられている。チンパンジーは、アロケアの程度が低い種として位置づけられ、したがって、こうした特徴はヒトがチンパンジーとの分岐後に獲得した形質ということになっている。
生活史上の特徴は、これ以外にも、言語進化・高い認知能力・共有志向性・二足での持久走などと、相互に複雑に関わっているとされる。こうした一連の議論の中では、チンパンジーや他の類人猿は「離乳が遅く、離乳すれば食物獲得に関しては独立し(幼少期がなく)、アロケアがない(協力的でない)」という形に要約されることになる。以下では、こうしたステレオタイプなイメージについて、チンパンジーの孤児の存在から再検討を加える。
タンザニア、マハレに生息するチンパンジーM集団では、1990年代前半までは孤児が多かったが、2000年代には孤児がほとんどいなくなった。このため、この間、孤児の観察をする機会自体が少なかったのだが、最近数頭の個体が母親を亡くし、孤児になった。これまでの記録を見ると、孤児で生存できた最低年齢は3歳である。アカンボウが乳首を吸うのを止めるのが4〜5歳くらいであることから、行動上の離乳と「本当の」離乳の時期とがずれている可能性が示唆される。3歳で生き残ることがあるとはいえ、より年長で母親を失った個体たちを見ても、やはり孤児は明らかに困っているように見える。つまり、裏を返せば、離乳後であっても母親の存在は大きいことが示唆される。
それでは、「母親の不在」は極限的な社会環境なのだろうか。先行研究では離乳後の孤児が無気力・食欲不振で3週間後に死亡した例が報告されており、母親への心理的依存が大きかったことが示唆されている。マハレでは、母親が隣接集団に一時的に移出してしまったコドモ雄が、その間攻撃を受ける頻度が増加した例が報告されている。また、孤児になると、ワカモノになった際の身体発達が遅れることがあるし、少なくとも雄では孤児になると期待余命まで生きられない傾向がある。行動的にも、孤児は不安の指標であるラフスクラッチが多く、遊びが少ないといった報告もある。
ならば、チンパンジーの孤児はどう生き延びるのだろうか。これまでの報告例などを総合すると、やはり誰か依存する相手が必要なようである。典型的なのは養子取りである。孤児がアカンボウの場合、養子取りが生じることがあり、この場合養母となる個体は、授乳以外のほとんどのケア(運搬・ベッドのシェアなど)を与える。もう少し年齢が高い孤児の場合、誰か特定のオトナ個体に追随することが多い。2015年に孤児になったテトという雌の場合、オトナ雄がよく毛づくろいをしてやっていたという印象がある。つまり、比較的年長の孤児にとっても、遊動の際に待ってくれたりかまってくれたりする存在がいることは重要である。
母親が生きている場合のアロケアは、かつて西田がマハレK集団を対象に調べて以来、定量的には調べられていない。ただし、いくつか印象的な観察例はある。たとえば、アカンボウが障害を持っており、母親が運搬に苦労していた際に、アカンボウの姉がかなりの時間面倒を見、その間に母親が樹上で食事をしていたといったものである。
チンパンジーは遊動生活を送るので、幼少個体にとって誰かに追随できることは重要である。アカンボウのうちは、母親にお腹や背中で運ばれ、その後、おおむね8〜9歳くらいまでは母親に追随する。つまり、すでに自力で食べることができるとしても、食物がある場所に辿り着くことに関しては母親に依存している。採食樹まで辿り着ければ、樹上ではアカンボウであっても、自由に動き、自分で食べることができる。母親もアカンボウを抱えている必要はなく、その間は採食に集中できる。一方、コドモ期の後半くらいになると、母親が食べていない時に自分で食べるということも多くなるが、母親はそれを待ってくれる。稀に母親が先に移動してしまい、迷子になると、7〜8歳の個体でも悲鳴を上げて母親を捜し回る。こうした事例からも遊動に関しての母親への依存が続いていることが示唆される。
こうした観察事例を見る限り、「ヒトでは離乳後のケアが必要で、類人猿ではそうではない」という現在の二項対立的な理解は極端であるように思える。チンパンジーでも、母親の必要性は離乳では終わらない。だからこそ、離乳後でも孤児になると不利益を被るのである。
チンパンジーにはアロケアがないと要約されるのは、頻度が少ないからであった。ただし、母親や子供の性格や相手によって、アロケアがどの程度生じるかどうかは異なる。母親が存命であれば、そもそもアロケアの必要性自体が低いのだが、いざ必要性が生じた場合(典型的には孤児になる場合)にはアロケアの重要性が増すだろう。孤児の存在は時代によってかなり偏りがある。どの時期にデータを取るかによっても養子取りやアロケアの量は変わるという点には注意が必要である。アロケアをする候補になる個体(兄・姉・子供のいない仲良しの雌など)がどの程度いるのかといった集団構成や社会的要因なども複雑に影響するだろう。
定住生活をするか、少なくともホームベースを持つヒトの場合、食物はそうした場所に持ち帰られることになる。一方、チンパンジーは食物の所に移動する。だから、ヒトの「ケア」が「食物を持ってくる」ことであるのに対して、チンパンジーの場合は「食物の所に連れていく」という形を取るのである。幼少期の定義も、こうしたことと関連して考える必要があるだろう。食物を「持ち帰る」という点ではたしかにヒトはチンパンジーと異なる。だが、離乳後にも子が食べるということに関してなんらかの「ケア」を与えるという点では共通している。ヒトだけが「ケア」を必要とする「幼少期」を持つといった形で断絶を強調する前に、それぞれの種の生活に応じた「ケア」のあり方をまずは考えるべきであろう。現在通説となっている「ヒトにしかない特徴」の中には、ヒト以外の霊長類の社会に、まったく同一の形ではないとしても、見られるものがまだまだあるはずである。
ヒトの授乳終了は、非産業社会の民族誌データなどから概ね2.5歳と言われており、一方、野生チンパンジーでは、平均5.2歳まで乳首接触が続く。第一大臼歯の萌出がヒトで6歳頃、チンパンジーで3歳頃であることを考えると、ヒトはかなり早い段階で離乳することになる。
これと連動したヒト特有の発達段階として「幼少期」が考えられている。これは、アカンボウ期とコドモ期の間に位置し、離乳と乳歯列の完成から6.9歳までとされている。幼少期には乳歯と小さな消化器系のため、噛むのが簡単で栄養が密な食物を食べる必要がある。自力では食物獲得ができないため、集団の年長メンバーが食物を集めて与えるということになる。すなわち、幼少期のヒトは自力で生きられず、親をはじめとした年長メンバーに栄養獲得を依存する必要がある。これを裏返せば、類人猿では、離乳さえしてしまえば自力で食物を得て生存できるということになる。
形質人類学では、「ワカモノ期」もまたヒトに特有であるとされる。その場合、ワカモノ期は、急激な成長スパートに特徴づけられる。チンパンジーではそうしたスパートが欠けているために、ヒトに特有であるとされているのだが、霊長類学者の多くは、ヒト以外の霊長類にもワカモノ期があると考えている。その場合は、性成熟から完全な体サイズになるまでといった定義になるが、以下ではワカモノ期についての検討は割愛する。
ヒトの女性は、閉経後(繁殖終了後)も長く生存する。この閉経後の期間が祖母期である。チンパンジーの雌の多くは死ぬまで出産を続けるため、これもまたヒトに特有の生活史段階ということになっている。祖母期の女性は、自分自身では繁殖をせず、自分の子の育児を手伝うことで、包括適応度を上げていると考えられている。幼少期の存在とも密接に関連し、母親が授乳を早く切り上げて、次の子の妊娠・出産にエネルギーを投資できるのも、祖母のような母親以外の養育者が協力するからだとされる。
こうした生活史上の特徴は、その他のさまざまなヒトの特徴と関連づけられて議論される。たとえば、脳が大型化し始めると、母乳だけでは成長に必要なタンパク質が不足する。このため、栄養を肉で補填する必要が生じ、早い段階での離乳が有利になるとする考え方がある。これは、肉食への移行とも関連する。肉の獲得については、かつては狩猟の重要性が強調されていたが、現在ではスキャベンジング(屍肉食)が重要であったと考える研究者が多い。スキャベンジングには、肉食獣が完全に去った後に、残った骨から骨髄などを利用するパッシブ・スキャベンジングと、より積極的に肉食獣を追い払って肉を獲得するパワー・スキャベンジングが考えられている。後者の場合、肉食獣と対峙するため、危険を伴い、若いオトナの死亡率が高くなると予測される。そうすると、子は早い段階で離乳しておいたほうが有利であることになる。
「超−協力(hyper-cooperation)」もまた、ヒトに独特な特徴であると考えられている。複数の霊長類種を対象にした実験で、アロケア(allocare:母以外による子へのケア)の存在と協力行動に連関があるとする結果がある。こうした結果をもとに、早い離乳と幼少期の存在によって協同育児(当然アロケアを伴う)が適応的になり、「超−協力」が進化したと考えられている。チンパンジーは、アロケアの程度が低い種として位置づけられ、したがって、こうした特徴はヒトがチンパンジーとの分岐後に獲得した形質ということになっている。
生活史上の特徴は、これ以外にも、言語進化・高い認知能力・共有志向性・二足での持久走などと、相互に複雑に関わっているとされる。こうした一連の議論の中では、チンパンジーや他の類人猿は「離乳が遅く、離乳すれば食物獲得に関しては独立し(幼少期がなく)、アロケアがない(協力的でない)」という形に要約されることになる。以下では、こうしたステレオタイプなイメージについて、チンパンジーの孤児の存在から再検討を加える。
タンザニア、マハレに生息するチンパンジーM集団では、1990年代前半までは孤児が多かったが、2000年代には孤児がほとんどいなくなった。このため、この間、孤児の観察をする機会自体が少なかったのだが、最近数頭の個体が母親を亡くし、孤児になった。これまでの記録を見ると、孤児で生存できた最低年齢は3歳である。アカンボウが乳首を吸うのを止めるのが4〜5歳くらいであることから、行動上の離乳と「本当の」離乳の時期とがずれている可能性が示唆される。3歳で生き残ることがあるとはいえ、より年長で母親を失った個体たちを見ても、やはり孤児は明らかに困っているように見える。つまり、裏を返せば、離乳後であっても母親の存在は大きいことが示唆される。
それでは、「母親の不在」は極限的な社会環境なのだろうか。先行研究では離乳後の孤児が無気力・食欲不振で3週間後に死亡した例が報告されており、母親への心理的依存が大きかったことが示唆されている。マハレでは、母親が隣接集団に一時的に移出してしまったコドモ雄が、その間攻撃を受ける頻度が増加した例が報告されている。また、孤児になると、ワカモノになった際の身体発達が遅れることがあるし、少なくとも雄では孤児になると期待余命まで生きられない傾向がある。行動的にも、孤児は不安の指標であるラフスクラッチが多く、遊びが少ないといった報告もある。
ならば、チンパンジーの孤児はどう生き延びるのだろうか。これまでの報告例などを総合すると、やはり誰か依存する相手が必要なようである。典型的なのは養子取りである。孤児がアカンボウの場合、養子取りが生じることがあり、この場合養母となる個体は、授乳以外のほとんどのケア(運搬・ベッドのシェアなど)を与える。もう少し年齢が高い孤児の場合、誰か特定のオトナ個体に追随することが多い。2015年に孤児になったテトという雌の場合、オトナ雄がよく毛づくろいをしてやっていたという印象がある。つまり、比較的年長の孤児にとっても、遊動の際に待ってくれたりかまってくれたりする存在がいることは重要である。
母親が生きている場合のアロケアは、かつて西田がマハレK集団を対象に調べて以来、定量的には調べられていない。ただし、いくつか印象的な観察例はある。たとえば、アカンボウが障害を持っており、母親が運搬に苦労していた際に、アカンボウの姉がかなりの時間面倒を見、その間に母親が樹上で食事をしていたといったものである。
チンパンジーは遊動生活を送るので、幼少個体にとって誰かに追随できることは重要である。アカンボウのうちは、母親にお腹や背中で運ばれ、その後、おおむね8〜9歳くらいまでは母親に追随する。つまり、すでに自力で食べることができるとしても、食物がある場所に辿り着くことに関しては母親に依存している。採食樹まで辿り着ければ、樹上ではアカンボウであっても、自由に動き、自分で食べることができる。母親もアカンボウを抱えている必要はなく、その間は採食に集中できる。一方、コドモ期の後半くらいになると、母親が食べていない時に自分で食べるということも多くなるが、母親はそれを待ってくれる。稀に母親が先に移動してしまい、迷子になると、7〜8歳の個体でも悲鳴を上げて母親を捜し回る。こうした事例からも遊動に関しての母親への依存が続いていることが示唆される。
こうした観察事例を見る限り、「ヒトでは離乳後のケアが必要で、類人猿ではそうではない」という現在の二項対立的な理解は極端であるように思える。チンパンジーでも、母親の必要性は離乳では終わらない。だからこそ、離乳後でも孤児になると不利益を被るのである。
チンパンジーにはアロケアがないと要約されるのは、頻度が少ないからであった。ただし、母親や子供の性格や相手によって、アロケアがどの程度生じるかどうかは異なる。母親が存命であれば、そもそもアロケアの必要性自体が低いのだが、いざ必要性が生じた場合(典型的には孤児になる場合)にはアロケアの重要性が増すだろう。孤児の存在は時代によってかなり偏りがある。どの時期にデータを取るかによっても養子取りやアロケアの量は変わるという点には注意が必要である。アロケアをする候補になる個体(兄・姉・子供のいない仲良しの雌など)がどの程度いるのかといった集団構成や社会的要因なども複雑に影響するだろう。
定住生活をするか、少なくともホームベースを持つヒトの場合、食物はそうした場所に持ち帰られることになる。一方、チンパンジーは食物の所に移動する。だから、ヒトの「ケア」が「食物を持ってくる」ことであるのに対して、チンパンジーの場合は「食物の所に連れていく」という形を取るのである。幼少期の定義も、こうしたことと関連して考える必要があるだろう。食物を「持ち帰る」という点ではたしかにヒトはチンパンジーと異なる。だが、離乳後にも子が食べるということに関してなんらかの「ケア」を与えるという点では共通している。ヒトだけが「ケア」を必要とする「幼少期」を持つといった形で断絶を強調する前に、それぞれの種の生活に応じた「ケア」のあり方をまずは考えるべきであろう。現在通説となっている「ヒトにしかない特徴」の中には、ヒト以外の霊長類の社会に、まったく同一の形ではないとしても、見られるものがまだまだあるはずである。
4.「霊長類の群れが生存できる極限の環境」(中川尚史)
1 はじめに
「ヒトはひとりで生きられない」、「ヒトは社会的な動物である」などと言われるように、ヒトが生きるということは社会的な存在として生きるということと同義である。ヒト以外の群居性の霊長類の場合、彼らにとって生きるということは基本的な社会単位であるところの「群れ」の一員として生きるということとほぼ 同義であるから、彼らが生存できる極限の環境を考える時、群れとして生存できる環境を考える必要がある。しかし、生存というとどうしても個体の生存、あるいは個体群の生存を考えることが多く、この少し考えれば当たり前のアプローチで霊長類の生存環境を捉えることは少なかった。
野生霊長類の研究において意識されることが少なかったもうひとつの問題に、時間的制約がある。ヒトは時間が有限であることを知っている。だからこそ、「時間がない、時間がない」と言って、睡眠時間を削り、食事の時間さえ惜しんで、時間に追われながら日々を送っている人は大勢いる。しかし野生の霊長類を観察していても、彼らが時間に追われていることを感じとれることはほとんどない。昼行性霊長類の場合、観察者の多くは日が暮れてから夜が明けるまでは彼らは眠っていると仮定しており、睡眠時間を削っていたとしても観察者は感じとりようがない。また、日中観察していても、じゅうぶんに休息し、毛づくろいに興じているように見えることが多いことも、霊長類研究者が時間を生存の制約として扱うことが少なかった理由であろう。
こうした状況の中、エチオピアにおいて河合雅雄らと同時期にゲラダヒヒの調査を行っていたイギリスの霊長類学者・人類学者であるロビン・ダンバーは、1992年『Time: a hidden constraint on the behavioral ecology of baboons』と題する論文を発表した。有名な『社会脳仮説』のもと論文となる『Neocortex size as a constraint on group size in primates』と同年の発表であることからも推察されるように、これらの2つの論文は相互に深く連関している。『社会脳仮説』は、霊長類の大脳新皮質率がそれぞれの種の平均的な群れサイズと正の相関をするという事実から、脳の進化を推し進めたのは社会の複雑さであることを唱えた説であるが、この関係をもとにヒトの大脳新皮質率からヒトの群れサイズを推定すると150人となり、狩猟採集民の平均的なバンドの大きさに一致するのだという。他方、ダンバーは、霊長類の毛づくろい時間が群れサイズと正の相関を示すことから、毛づくろいは群れの紐帯を維持するためのいわば膠(にかわ)の役割を果たしていると考えている。ここで今度は、150人といういわばヒトの群れサイズからヒトの毛づくろい時間を逆に推測すると時間が足りなくなる計算になり、群れの紐帯を維持すべく同時に多くの個体と交わすことのできる手段として言語を編み出したという説を提唱するに至った。さて、ヒトを含め霊長類では、群れの大きさによって決まる毛づくろい時間のほかに、採食や移動という生きていく上で必須の活動があり、それらの時間割合は気温や降水量などの環境要因によって決まるため、生息環境によって、一定の群れサイズを維持するために時間が制限要因になるという発想に至った。
本稿では、ニホンザルが個体として、さらには群れで生きていく上で時間が制限要因になっていることを示唆する私自身の研究を紹介したあと、ダンバーによる上述の研究、ならびにその発展型を紹介することで、霊長類が群れで生存できる極限の環境について考えてみる。
2 活動時間配分の決定要因
金華山、ならびに屋久島に生息するニホンザルの活動時間配分を年間通じて調査したところ、いずれの地域も移動と休息は概ね採食と逆相関する、日長と相関する月変動を示すが、毛づくろいは連動しないことが分かった。他方、地域間比較を通じて、金華山は屋久島に比べ採食時間がかなり長い代わりに、毛づくろい時間がかなり短かった。これは、金華山は採食速度が低いことに加え、低温のため体温維持エネルギーが高いためと考えられた。こうした結果は、ヒヒの地域間比較研究と一致する。
こうした結果、ならびにヒヒの地域間比較の研究から、以下のような一般論が導かれた。1)気象要因(気温や降水量)、あるいはそれと連動した食物の量や質の季節的・地域的変動と連関して、採食時間や移動時間は増減する。2)1日の活動可能時間には限界があるため、休息時間が採食時間や移動時間の増加分を吸収する形で減少する。3)それに対し毛づくろいは、季節的にはその時間長が保持される傾向にある。4)しかしながら、地域的には、1)と2)による毎年繰り返される休息時間の逼迫が、毛づくろい時間をわずかながら減少させる。5)毛づくろい時間が相関を示すのは、群れサイズとであり、毛づくろいが群れの紐帯を維持する膠の役割を果たしていることを示唆している。
3 ロビン・ダンバーの生態学的に許容できる最大の群れサイズ(Maximum ecologically tolerable group size)
3-1 Dunbar (1992) の旧モデル
アヌビスヒヒ、キイロヒヒ、チャクマヒヒ、ギニアヒヒ、マントヒヒの全4亜種のヒヒ種の行動時間配分や群れサイズ、生息地における環境要因を調べ上げ、各活動時間割合をさまざまな環境要因や群れサイズで表す最適モデルを求めた。その後、採食時間および移動時間については、降水量と気温という環境要因、ならびに群れサイズだけの関係に集約させ、毛づくろい時間については以下のような関係式を求めた。
S(毛づくろい時間)= 4.533 + 0.0764N(群れサイズ)
降水量と気温と群れサイズから採食時間や移動時間を推定し、休息時間については5%だけ残して残りの時間を毛づくろい時間に回せると仮定し、その地域で生態学的に許容できる最大の群れサイズ(Maximum ecologically tolerable group size)を求めた。その結果、年平均気温が10℃以下、あるいは35℃以上である場所や、年間降水量が2700mm以上だと気温が30℃以上でないと、群れを形成することができないこと。また、あまり降水量が少ないと大きな群れを形成できないこと。つまり、中程度の気温と降水量で、大きな群れが形成できるという推定結果が得られた。
そこで、以下の式から計算できる生態ストレス度を算出した。
生態ストレス度=群れサイズ(観察値)/降水量と気温から予測されたその地域で許容できる最大群れサイズ
そのうえで、採食時間ならびに[休息ならびに毛づくろいの合計時間]を、生態ストレス度が1以上である群れと1未満である群れを比較したところ、採食には大きな差は認められなかったが、[休息ならびに毛づくろいの合計時間]は、1以上である群れが1未満である群れより非常に短かった。この結果は、ストレス度が1より高いと[休息と毛づくろい時間]を減らして採食時間を維持していることを示している。さらに、採食中、頻繁に分派している群れと、そうでない群れのストレス度を比較したところ、前者でストレス度が高いことも分かった。
3-2 Bettridge、 Lehmann、 and Dunbar (2010)の新モデル
Dunbar (1992)以後のデータを加え、次の3つの仮定、1)採食時間と移動時間は独立に気候要因、食物、そして群れサイズで決まると仮定、2)S(毛づくろい時間)= 1.55 + 0.23N(群れサイズ)、3)毛づくろい時間割合は最低1%、その他の時間配分はすべてで5%が最低限必要、に基づき、群れサイズ1頭から始まり、すべての活動時間配分の合計が100%になるまで1頭ずつ増やしていくことにより、その地域で生態学的に許容できる最大の群れサイズを求めた。その結果、大ざっぱに言えば、乾燥が進むと採食時間が減り(ただし、完全に砂漠になり降水量変動少ないと増える要素もある)、気温変動が激しい分休息時間が増え、年平均気温が上がる分休息時間が増え(体温上昇回避)、移動時間は減少するという推定結果が得られた。しかし、旧モデルと比べ、変数が複雑で解釈し辛くなった点が認められた。
4 最小の群れサイズ(Mimimum permissible group size)
その地域の最小の群れサイズは、捕食の危険度で決まるという仮説のもと、捕食者の種類、有無ごとに実際の最小群れサイズを比較した。捕食者がいる場所のほうが最小群れサイズが大きく、その効果はヒョウよりライオンが大きいこと、捕食者密度が高いほど最小群れサイズ大きいこと、オトナメスの体重が重いほど最小群れサイズ小さいことなど、仮説を支持する結果が得られた。
5 生態学的に許容できる最大の群れサイズの予測値と実測値
新モデルで求められた生態学的に許容できる最大の群れサイズの予測値を、平均群れサイズの実測値を比較したところ、概ね実測値は小さかった。また、最大群れサイズの実測値を比較したところ、概ね一致する傾向にあった。ただし、いずれの場合も、マントヒヒは、実測値のほうがかなり高いはずれ値となった。
6 実現ニッチ空間(Realisable niche space)
実現ニッチ空間にあると予測される地域と実際のヒヒの分布は、概ね一致した。また、ヒヒがいない場所と実際にいる場所の環境要因から予測される活動時間配分を比較すると、採食時間のみ大きな差が認められ、前者で非常に長いという結果が得られた。この結果は、ヒヒが採食時間の要求が非常に高い生息地では住めないことを示している。
7 まとめ
捕食の危険が最小群れサイズの決定に働くという点には新奇性は特にないが、群れメンバー間の紐帯維持に働くため群れサイズによって決まる毛づくろい時間、ならびに気温や降水量などの環境要因によって決まる採食時間や移動時間を合わせた活動の時間的制約により、それぞれの環境できる最大の群れサイズが決まるという着想は、新奇性が非常に高い。また、本発表で紹介したヒヒ属のみならず、チンパンジー属、クモザル属、アカコロブス属、シロクロコロブス属、サバンナモンキー属、ゴリラ属でも同様の分析を行い、最終的にはいずれも実際の分布との一致度が高いという結果が得られていることは、本モデルの汎用性、頑強性の高さを示している。
しかしながら、幾つかの懸念がないわけではない。1)ヒヒの新モデルでみられたように変数が複雑で結果の解釈がイメージしづらい場合があること。2)私のニホンザルの地域間比較では、採食時間が限界まで長くなる直接的理由が、採食速度が低いこと、気温が低いことと明確であったのに比べて、必ずしも明瞭でないこと。多雪地のニホンザルでは、食物の量が少なく低質となると、採食も移動も極端に減らしてエネルギーを消費する戦術に転ずることが知られており、このように時間が制約とはならないこともある。3)実際の分布は、一般的には環境要因だけでなく、地史や種間関係が効いていると考えられること。
8 おわりに
「ひとりで生きられない」ヒトが、農山間部では過疎化により集落の呈をなさなくなり、限界集落と呼ばれる状況になっている。昨今、その農山間部の大きな問題であるニホンジカ、イノシシ、ニホンザルを主とした田畑や果樹園の作物を荒らす獣害は、究極的には限界集落問題に行き着くと言われている。過疎化が耕作放棄地を生むとともに獣を誘因し、少数の高齢者だけでは、作物を守り切れず、栄養価の高い作物が獣の増加を招くという悪循環である。これらの獣はヒトにとって捕食者ではなくいわば競争者であるが、ダンバーモデルの捕食の危険の高さで群れサイズの下限が決まるということのアナロジーとして捉えることができよう。
他方、毛づくろい時間がじゅうぶんとれないことが群れサイズの上限を決めるという発想は、逆に過密に悩まされる大都市の空虚な人間関係を連想させる。睡眠時間を削り、食事の時間さえ惜しんで働き、忙しさを口実に人づきあいもせず、隣近所にさえ無関心である。同性間の人づきあいはおろか、最近は異性間の人づきあいもしなくなっているという。大都市ではますます晩婚化が進み、しかしそれでも大都市への人口流入は止まらず、こちらでも悪循環に陥っているとも言える。
【参考文献】
「ヒトはひとりで生きられない」、「ヒトは社会的な動物である」などと言われるように、ヒトが生きるということは社会的な存在として生きるということと同義である。ヒト以外の群居性の霊長類の場合、彼らにとって生きるということは基本的な社会単位であるところの「群れ」の一員として生きるということと
野生霊長類の研究において意識されることが少なかったもうひとつの問題に、時間的制約がある。ヒトは時間が有限であることを知っている。だからこそ、「時間がない、時間がない」と言って、睡眠時間を削り、食事の時間さえ惜しんで、時間に追われながら日々を送っている人は大勢いる。しかし野生の霊長類を観察していても、彼らが時間に追われていることを感じとれることはほとんどない。昼行性霊長類の場合、観察者の多くは日が暮れてから夜が明けるまでは彼らは眠っていると仮定しており、睡眠時間を削っていたとしても観察者は感じとりようがない。また、日中観察していても、じゅうぶんに休息し、毛づくろいに興じているように見えることが多いことも、霊長類研究者が時間を生存の制約として扱うことが少なかった理由であろう。
こうした状況の中、エチオピアにおいて河合雅雄らと同時期にゲラダヒヒの調査を行っていたイギリスの霊長類学者・人類学者であるロビン・ダンバーは、1992年『Time: a hidden constraint on the behavioral ecology of baboons』と題する論文を発表した。有名な『社会脳仮説』のもと論文となる『Neocortex size as a constraint on group size in primates』と同年の発表であることからも推察されるように、これらの2つの論文は相互に深く連関している。『社会脳仮説』は、霊長類の大脳新皮質率がそれぞれの種の平均的な群れサイズと正の相関をするという事実から、脳の進化を推し進めたのは社会の複雑さであることを唱えた説であるが、この関係をもとにヒトの大脳新皮質率からヒトの群れサイズを推定すると150人となり、狩猟採集民の平均的なバンドの大きさに一致するのだという。他方、ダンバーは、霊長類の毛づくろい時間が群れサイズと正の相関を示すことから、毛づくろいは群れの紐帯を維持するためのいわば膠(にかわ)の役割を果たしていると考えている。ここで今度は、150人といういわばヒトの群れサイズからヒトの毛づくろい時間を逆に推測すると時間が足りなくなる計算になり、群れの紐帯を維持すべく同時に多くの個体と交わすことのできる手段として言語を編み出したという説を提唱するに至った。さて、ヒトを含め霊長類では、群れの大きさによって決まる毛づくろい時間のほかに、採食や移動という生きていく上で必須の活動があり、それらの時間割合は気温や降水量などの環境要因によって決まるため、生息環境によって、一定の群れサイズを維持するために時間が制限要因になるという発想に至った。
本稿では、ニホンザルが個体として、さらには群れで生きていく上で時間が制限要因になっていることを示唆する私自身の研究を紹介したあと、ダンバーによる上述の研究、ならびにその発展型を紹介することで、霊長類が群れで生存できる極限の環境について考えてみる。
2 活動時間配分の決定要因
金華山、ならびに屋久島に生息するニホンザルの活動時間配分を年間通じて調査したところ、いずれの地域も移動と休息は概ね採食と逆相関する、日長と相関する月変動を示すが、毛づくろいは連動しないことが分かった。他方、地域間比較を通じて、金華山は屋久島に比べ採食時間がかなり長い代わりに、毛づくろい時間がかなり短かった。これは、金華山は採食速度が低いことに加え、低温のため体温維持エネルギーが高いためと考えられた。こうした結果は、ヒヒの地域間比較研究と一致する。
こうした結果、ならびにヒヒの地域間比較の研究から、以下のような一般論が導かれた。1)気象要因(気温や降水量)、あるいはそれと連動した食物の量や質の季節的・地域的変動と連関して、採食時間や移動時間は増減する。2)1日の活動可能時間には限界があるため、休息時間が採食時間や移動時間の増加分を吸収する形で減少する。3)それに対し毛づくろいは、季節的にはその時間長が保持される傾向にある。4)しかしながら、地域的には、1)と2)による毎年繰り返される休息時間の逼迫が、毛づくろい時間をわずかながら減少させる。5)毛づくろい時間が相関を示すのは、群れサイズとであり、毛づくろいが群れの紐帯を維持する膠の役割を果たしていることを示唆している。
3 ロビン・ダンバーの生態学的に許容できる最大の群れサイズ(Maximum ecologically tolerable group size)
3-1 Dunbar (1992) の旧モデル
アヌビスヒヒ、キイロヒヒ、チャクマヒヒ、ギニアヒヒ、マントヒヒの全4亜種のヒヒ種の行動時間配分や群れサイズ、生息地における環境要因を調べ上げ、各活動時間割合をさまざまな環境要因や群れサイズで表す最適モデルを求めた。その後、採食時間および移動時間については、降水量と気温という環境要因、ならびに群れサイズだけの関係に集約させ、毛づくろい時間については以下のような関係式を求めた。
S(毛づくろい時間)= 4.533 + 0.0764N(群れサイズ)
降水量と気温と群れサイズから採食時間や移動時間を推定し、休息時間については5%だけ残して残りの時間を毛づくろい時間に回せると仮定し、その地域で生態学的に許容できる最大の群れサイズ(Maximum ecologically tolerable group size)を求めた。その結果、年平均気温が10℃以下、あるいは35℃以上である場所や、年間降水量が2700mm以上だと気温が30℃以上でないと、群れを形成することができないこと。また、あまり降水量が少ないと大きな群れを形成できないこと。つまり、中程度の気温と降水量で、大きな群れが形成できるという推定結果が得られた。
そこで、以下の式から計算できる生態ストレス度を算出した。
生態ストレス度=群れサイズ(観察値)/降水量と気温から予測されたその地域で許容できる最大群れサイズ
そのうえで、採食時間ならびに[休息ならびに毛づくろいの合計時間]を、生態ストレス度が1以上である群れと1未満である群れを比較したところ、採食には大きな差は認められなかったが、[休息ならびに毛づくろいの合計時間]は、1以上である群れが1未満である群れより非常に短かった。この結果は、ストレス度が1より高いと[休息と毛づくろい時間]を減らして採食時間を維持していることを示している。さらに、採食中、頻繁に分派している群れと、そうでない群れのストレス度を比較したところ、前者でストレス度が高いことも分かった。
3-2 Bettridge、 Lehmann、 and Dunbar (2010)の新モデル
Dunbar (1992)以後のデータを加え、次の3つの仮定、1)採食時間と移動時間は独立に気候要因、食物、そして群れサイズで決まると仮定、2)S(毛づくろい時間)= 1.55 + 0.23N(群れサイズ)、3)毛づくろい時間割合は最低1%、その他の時間配分はすべてで5%が最低限必要、に基づき、群れサイズ1頭から始まり、すべての活動時間配分の合計が100%になるまで1頭ずつ増やしていくことにより、その地域で生態学的に許容できる最大の群れサイズを求めた。その結果、大ざっぱに言えば、乾燥が進むと採食時間が減り(ただし、完全に砂漠になり降水量変動少ないと増える要素もある)、気温変動が激しい分休息時間が増え、年平均気温が上がる分休息時間が増え(体温上昇回避)、移動時間は減少するという推定結果が得られた。しかし、旧モデルと比べ、変数が複雑で解釈し辛くなった点が認められた。
4 最小の群れサイズ(Mimimum permissible group size)
その地域の最小の群れサイズは、捕食の危険度で決まるという仮説のもと、捕食者の種類、有無ごとに実際の最小群れサイズを比較した。捕食者がいる場所のほうが最小群れサイズが大きく、その効果はヒョウよりライオンが大きいこと、捕食者密度が高いほど最小群れサイズ大きいこと、オトナメスの体重が重いほど最小群れサイズ小さいことなど、仮説を支持する結果が得られた。
5 生態学的に許容できる最大の群れサイズの予測値と実測値
新モデルで求められた生態学的に許容できる最大の群れサイズの予測値を、平均群れサイズの実測値を比較したところ、概ね実測値は小さかった。また、最大群れサイズの実測値を比較したところ、概ね一致する傾向にあった。ただし、いずれの場合も、マントヒヒは、実測値のほうがかなり高いはずれ値となった。
6 実現ニッチ空間(Realisable niche space)
実現ニッチ空間にあると予測される地域と実際のヒヒの分布は、概ね一致した。また、ヒヒがいない場所と実際にいる場所の環境要因から予測される活動時間配分を比較すると、採食時間のみ大きな差が認められ、前者で非常に長いという結果が得られた。この結果は、ヒヒが採食時間の要求が非常に高い生息地では住めないことを示している。
7 まとめ
捕食の危険が最小群れサイズの決定に働くという点には新奇性は特にないが、群れメンバー間の紐帯維持に働くため群れサイズによって決まる毛づくろい時間、ならびに気温や降水量などの環境要因によって決まる採食時間や移動時間を合わせた活動の時間的制約により、それぞれの環境できる最大の群れサイズが決まるという着想は、新奇性が非常に高い。また、本発表で紹介したヒヒ属のみならず、チンパンジー属、クモザル属、アカコロブス属、シロクロコロブス属、サバンナモンキー属、ゴリラ属でも同様の分析を行い、最終的にはいずれも実際の分布との一致度が高いという結果が得られていることは、本モデルの汎用性、頑強性の高さを示している。
しかしながら、幾つかの懸念がないわけではない。1)ヒヒの新モデルでみられたように変数が複雑で結果の解釈がイメージしづらい場合があること。2)私のニホンザルの地域間比較では、採食時間が限界まで長くなる直接的理由が、採食速度が低いこと、気温が低いことと明確であったのに比べて、必ずしも明瞭でないこと。多雪地のニホンザルでは、食物の量が少なく低質となると、採食も移動も極端に減らしてエネルギーを消費する戦術に転ずることが知られており、このように時間が制約とはならないこともある。3)実際の分布は、一般的には環境要因だけでなく、地史や種間関係が効いていると考えられること。
8 おわりに
「ひとりで生きられない」ヒトが、農山間部では過疎化により集落の呈をなさなくなり、限界集落と呼ばれる状況になっている。昨今、その農山間部の大きな問題であるニホンジカ、イノシシ、ニホンザルを主とした田畑や果樹園の作物を荒らす獣害は、究極的には限界集落問題に行き着くと言われている。過疎化が耕作放棄地を生むとともに獣を誘因し、少数の高齢者だけでは、作物を守り切れず、栄養価の高い作物が獣の増加を招くという悪循環である。これらの獣はヒトにとって捕食者ではなくいわば競争者であるが、ダンバーモデルの捕食の危険の高さで群れサイズの下限が決まるということのアナロジーとして捉えることができよう。
他方、毛づくろい時間がじゅうぶんとれないことが群れサイズの上限を決めるという発想は、逆に過密に悩まされる大都市の空虚な人間関係を連想させる。睡眠時間を削り、食事の時間さえ惜しんで働き、忙しさを口実に人づきあいもせず、隣近所にさえ無関心である。同性間の人づきあいはおろか、最近は異性間の人づきあいもしなくなっているという。大都市ではますます晩婚化が進み、しかしそれでも大都市への人口流入は止まらず、こちらでも悪循環に陥っているとも言える。
【参考文献】
- Agetsuma, N., and N. Nakagawa. 1998. Effects of habitat differences on feeding behaviors of Japanese monkeys: Comparison between Yakushima and Kinkazan. Primates 39: 275-289.
- Bettridge, C. M., and R. I. M. Dunbar. 2012. Predation as a determinant of minimum group size in baboons. Folia primatologica 83: 332-352.
- Bettridge, C., J. Lehmann, and R. I. M. Dunbar. 2010. Trade-offs between time, predation risk and life history, and their implications for biogeography: a systems modelling approach with a primate case study. Ecological Modelling 221: 777-790.
- Dunbar, R. I. M. 1992. Time: a hidden constraint on the behavioral ecology of baboons. Behavioral Ecology and Sociobiology 31: 35-49.
- Dunbar, R. I. M., A. H. Korstjens, and J. Lehmann. 2009. Time as an ecological constraint. Biological Reviews of the Cambridge Philosophical Society 84: 413-429.
- Dunbar, R. I. M., and M. Sharman. 1984. Is social grooming altruistic? Zeitschrift Fur Tierpsychologie 64: 163-173.
- 伊谷純一郎・寺嶋秀明.2001. アフリカの植生図.一試案の提示.人間文化 15: 15-18.
- Korstjens, A. H., and R. I. M. Dunbar. 2007. Time constraints limit group sizes and distribution in red and black-and-white Colobus. International Journal of Primatology 28: 551-575.
- Korstjens, A. H., I. L. Verhoeckx, and R. I. M. Dunbar. 2006. Time as a constraint on group size in spider monkeys. Behavioral Ecology and Sociobiology 60: 683-694.
- Lehmann, J., A. H. Korstjens, and R. I. M. Dunbar. 2007. Group size, grooming and social cohesion in primates. Animal Behaviour 74: 1617-1629.
- Lehmann, J., A. H. Korstjens, and R. I. M. Dunbar. 2008. Time management in great apes: implications for gorilla biogeography. Evolutionary Ecology Research 10: 517-536.
- 中川尚史.1999.『食べる速さの生態学―サルたちの採食戦略』,京都大学学術出版会,京都.
- Willems, E. P., and R. A. Hill. 2009. A critical assessment of two species distribution models: a case study of the vervet monkey (Cercopithecus aethiops). Journal of Biogeography 36: 2300-2312.
5.「牧畜民の遊動再考ー諸環境との相互作用から」(河合香吏)
1.はじめに
本共同研究課題(「人類社会の進化史的基盤研究」)の第4期は「生存・環境・極限」をテーマに進められてきたが,「極限」については分析に耐えうるフィールドデータを持ちあわせていないため,本発表では主として「生存」と「環境」に着目した。まず「環境」を次のようにとらえる。「人間を含むあらゆる生物は単体としてただそれだけで存在することはできず,その生物個体を取り囲む環境の諸要素と相互作用しつつ生きている。…人間の社会性の進化を考えるとき,主体にとってもっとも重要な環境要素となるのは同種の他個体の存在である」(足立2016:358)。また,「場」という概念について,船曳は「人間は,出会い,対面する他の人間と相互に了解の関係を取ることが可能であり(2009:294),その関係の可能性を保っている,無限の位置の広がりを「場」と呼ぼう(2016:420)」と提唱し,足立もまた,生態学における「場」の理論に触れながら,「「場」とは生活する生物の個体間での相互作用が不断に接続する場所であり…そこに生息する生物の行為が,無機的な環境や同種異種の他個体の行動と相互作用を続けている限りにおいて,「場」が成立する(足立2013:275–280)」と指摘する。こうした意味で「場」という語を用いる。
ヒトは群居性という形質を生物学的=進化的基盤としてもつ霊長類の一員である。群れ=集団で暮らすのだから,「生存」には何がしかのかたちで「他者(他個体)」が介在するはずで,「生存」は社会的なもの,すなわち,文脈依存的,言いかえれば,非決定論的で可変的(フレキシブル)でコミュニケーショナルなものとなるだろう。ヒトはこうした社会性socialityを基盤に,どのような環境に,どのように生存しているのか。生存の「場」としての環境を,(1)自然生態的環境,(2)社会的環境,(3)象徴的環境に分析上区別して,牧畜民の遊動生活を材料として考えた。
2.ドドスの遊動生活
牧畜民の遊動生活は,植生が貧弱で水の乏しい乾燥地という厳しい自然環境を効率よく利用するための生態学的適応として分析される一方で,人間関係の葛藤や儀礼の開催,隣接集団との「戦い」等の社会的,政治的,宗教的な要因の関与が指摘されてきた。
ドドスはウガンダ北東部のケニア,スーダンとの三国国境地域に住むウシ牧畜民である。生産・消費の単位は父系の拡大家族であり,集落(ere)と家畜キャンプ(awi)に家族のメンバーと保有家畜が分散し,それぞれに自律的に日常生活が営まれている。集落は半定住的であるが,家畜キャンプは数日から1〜2カ月程度の間隔で頻繁に移動を繰り返し,移動距離も数キロメートルから100キロメートルを超える場合もある。したがって,ドドスの遊動生活を形作っているのは実質的には家畜キャンプの移動と言って差し支えない。
3. キャンプ移動の契機
3-1 家畜キャンプの移動の事例
移動の概要をつかむため,2003年6月2日〜7月2日における家畜キャンプ移動の事例を紹介した(表1,河合 2006:185)。
表1 家畜群の移動の経緯(2003.6.2-7.2)
この期間の家畜キャンプの移動は,形の上ではモルアンガピオンを基点に,モルエセとロクワカラモエの2カ所をそれぞれ往復したうえでロイタへ移動しているが,これらの移動の契機となったのは,エンゲレイの摂取が家畜飼育に関与する生態的な理由であることを除けば,預言者の指示や腸占いの啓示を直接的な契機としていた。こうした具体例を踏まえ,移動の契機となったのはどのような環境の変化なのか,ドドスの家畜キャンプ移動が3つの質的に異なる環境との相互作用としてあらわれる様子を追った。
3-2 自然生態的環境
上記の事例にはなかったが,主として草や水の時空間配置の変化に基づくキャンプ移動がある。また,事例のように家畜の成育に不可欠とされている塩分を含む草や土を定期的に採食させるためにキャンプ移動をすることがある。移動の契機となるこれらの要素を含む環境を「自然生態的環境」と呼ぶ。
ドドスにおける自然生態的環境は,降水量の少ない乾燥・半乾燥地における不規則で不安定で予測の難しい気まぐれな気象条件のもとにあり,しばしば旱魃に見舞われる。また,年間通じてほとんど雨の降らない大旱魃に襲われることもある。他方,大雨や洪水も問題となっており(2000年以降に目立つようになった。エルニーニョなど地球規模の気象変動,地球温暖化の影響による現象か?),鉄砲水で家畜が溺死したり,牧草地が長期間水浸しになったり泥水が流れ込むことによって牧草が生えなくなる(根腐れ?)等の被害がある。
自然生態的環境への適応が果たされないと家畜群の維持に支障を来し,ひいては人間の生存が脅かされる。このことを人びとはよく認識しており,自然生態的環境の変化(劣化)への「気づき」により,緊急に対処法が話し合われ,キャンプ移動が決行される。
そもそも,乾燥地という,そのままでは利用できない土地を,家畜を媒介することによって,人間の生息域としてきた乾燥地適応としての牧畜という生業の成立に関わる位置づけは普遍的なものである。だが,「水と草を求めて遊動する牧畜民」というイメージは素朴であり,現実的ではない。自然生態的環境の条件からは移動する必要がないのに移動したり,移動先の条件が自然生態的環境としては厳しすぎるにもかかわらず,あえて困難な場所へ移動したりすることもあるからである。事例では6回の移動の内3回が預言者の指示や腸占いの結果(啓示)がキャンプ移動の契機となっていた。次に述べる社会的環境や象徴的環境への対処を優先する事態が頻繁に起こっているのである。
3-3 社会的環境
ヒトが集団で生きる以上,環境は他者との相互作用の「場」である。そうした他者=同種他個体との相互作用の働く「場」を「社会的環境」と呼ぶ。社会的環境の要素である他者の作用によるキャンプ移動として,自民族内で放牧地を集中させない等の調整は常に意識的にされているが,隣接する他民族集団との関係もキャンプ移動に強く作用する。他集団のレイディングを避けるための移動が実に頻繁なのである。
「(家畜キャンプは)一カ所に長くとどまらない」という言説はよく耳にするが,その理由としては,「草や水が不足する」といった自然生態的環境の諸要素の作用よりもむしろ,「長く滞在すると家畜の居場所や放牧ルートが敵に知られ,放牧中の待ち伏せやキャンプ狙いの襲撃といったレイディングの危険が増すからだ」といった説明の方がより多く聞かれた。キャンプ移動は,自然生態的環境の要素である地勢や植生,採食地や給水地の状態等,移動先が放牧地やキャンプ設営地として適当かどうかの条件を満たしつつ,レイディングへの警戒とも不可分である。レイディングは牧畜民の生業基盤である家畜を群れごと奪われる,人びとの生存に直接的に関わる重大事であり,さまざまな方法でこの危険に対処し,また,キャンプ移動が決行されている。
社会的環境を形作る言説情報や目撃情報とともに,預言者の言葉や腸占いの啓示はレイディングの危険を「知る」よすがとなっていた。それは「兆し」「予兆」「気づき」といったものでもあるが,これらをもとに家畜を守るための行為が選ばれ,実践される。これについて次に述べる。
3-4 象徴的環境
自然生態的環境,社会的環境に加えて,ヒトには「いま,ここ」ではない,時空間を越えて,想像力によって創造される環境がある。これを「象徴的環境」と呼ぶ。ドドスにおいて象徴的環境を作り上げている諸要素の筆頭は「啓示」である。啓示は,主として,(1)預言者(呪医emuron)による夢見やサンダル占い,(2)腸占い,(3)個人的な夢見やサンダル占いなどの結果としてもたらされるが,象徴的環境の要素として日常的であり,かつ人びとが絶大な信頼を置くのは「腸占い」である。キャンプ移動に,腸占いが強くかかわる事例には事欠かない。
「啓示」は,近い将来の出来事(主として危険)を示唆するが,それは現実味を帯びた事態=現実問題として,人びとはさまざまな方法でこれに対処する。家畜の供犠を伴う大規模な儀礼の開催,特定の場所への呪物の設置,まじない的な儀礼的行為等とともに,日帰り放牧のルート変更や家畜キャンプの移動がここに含まれる。象徴的環境の諸要素は,「草がなくなった」「井戸の水が涸れた」などの自然生態的環境の要素の変化と同じ地平の出来事として作用するし,どこそこでレイディングがあったといった言説情報や足跡などの痕跡や人影などの目撃情報によるレイディングの「兆し」といった社会的環境を補強する要素として,人々の活動/行為選択に作用する。象徴的環境の諸要素は,「(生物学的)生存」にとってはしばしば明らかに過剰な行為である。だが,それは,人びとにとっては「生存」に直結する現実的な課題として迫って(作用して)くる事態であり,絶対に何らかの対処をすべき作用である。象徴的環境は想像力によって構築される世界だが,現実の物理空間を「超越」しておらず ,しばしば象徴的ではない,生業活動や日常生活上の諸行為を導き出すという点で,ドドスにとっては現実な日常世界と地続きのものとしてある。
3-5 レイヤーとしての環境
以上,3つの環境は,それぞれに独立しているのではないことは明らかである。また,「遊動」という生活様式にとって,どれがより優越しているわけでもない。地理空間上にある特定の場所は,自然生態的環境としてあるとともに,そこは社会的環境が投影された場所でもあり,象徴的環境として意味づけられた場所でもある。ドドスの遊動生活は,そうした質の異なる3つの環境のレイヤー構造=重層的な「場」に展開されるのであり,それぞれの環境からの作用を受けながら,それに対処するように,つまり3つの環境との間の相互作用として現れたものとしてある。
4. 環境の要素としての他者(他民族集団):その存在様態
キャンプ移動の契機となった他者はしばしば「情報」によって生成する存在であった。ここで言う情報は,(1)言説情報や目撃情報および(2)夢見や腸占いによる「啓示」という2種類が区別できる。前者は社会的環境の要素としての他者,後者は象徴的環境の要素としての他者という異なる2つの他者の存在様態を生成する。だが,この2つを人びとが異なる存在様態としてとらえているかどうかは不明である。これら2つが意味をもつのは,他者がレイディングを仕掛けてくる事態に際してであり,これに対処するためにパトロールの強化,儀礼の開催,呪物の設置,放牧コースの変更等をし,最終的には家畜キャンプの移動が選択される。牧畜民の遊動生活は自然生態的環境への適応であるとともに,隣接してともに生きる他者と相互作用する社会的環境や象徴的環境に大きく左右されていたのである。
こうした環境世界のあり方は,ヒトに特異的か?トマセロ(2013:53)によれば,「ヒトは他の類人猿のような物理的・社会的世界を生きているだけでなく,自分たち自身で作りあげた制度的・文化的世界…を生きている」。だが,これまでの共同研究を通じて制度的・文化的世界も萌芽的であれ霊長類にも認められることをわれわれは明らかにしてきた。人間は言葉を持つが故に,言葉が前面に出てしまうが,言葉を使う以前に未来についての「兆し」「予兆」「気づき」は,環境世界の諸要素の作用として既にあるのではないか。そのとき近い未来を「知る」術は霊長類の環境との相互作用と異なるといえるのか。検討の余地がある。
5.おわりに
結論めいたことを言う準備はできなかったが,一連の共同研究を通じて考えようとしてきたのは,隣接する民族集団が「敵対しつつも共存している」状況をいかに理解できるかであり,それは「他者とともに生きる」術(=高度な社会性という能力)の進化史的基盤を問うものであった。本発表で検討してきた「環境」の生成,成立とその利用がそれといかに関わるのかという問いは,今後の課題としたい。
【引用文献】
本共同研究課題(「人類社会の進化史的基盤研究」)の第4期は「生存・環境・極限」をテーマに進められてきたが,「極限」については分析に耐えうるフィールドデータを持ちあわせていないため,本発表では主として「生存」と「環境」に着目した。まず「環境」を次のようにとらえる。「人間を含むあらゆる生物は単体としてただそれだけで存在することはできず,その生物個体を取り囲む環境の諸要素と相互作用しつつ生きている。…人間の社会性の進化を考えるとき,主体にとってもっとも重要な環境要素となるのは同種の他個体の存在である」(足立2016:358)。また,「場」という概念について,船曳は「人間は,出会い,対面する他の人間と相互に了解の関係を取ることが可能であり(2009:294),その関係の可能性を保っている,無限の位置の広がりを「場」と呼ぼう(2016:420)」と提唱し,足立もまた,生態学における「場」の理論に触れながら,「「場」とは生活する生物の個体間での相互作用が不断に接続する場所であり…そこに生息する生物の行為が,無機的な環境や同種異種の他個体の行動と相互作用を続けている限りにおいて,「場」が成立する(足立2013:275–280)」と指摘する。こうした意味で「場」という語を用いる。
ヒトは群居性という形質を生物学的=進化的基盤としてもつ霊長類の一員である。群れ=集団で暮らすのだから,「生存」には何がしかのかたちで「他者(他個体)」が介在するはずで,「生存」は社会的なもの,すなわち,文脈依存的,言いかえれば,非決定論的で可変的(フレキシブル)でコミュニケーショナルなものとなるだろう。ヒトはこうした社会性socialityを基盤に,どのような環境に,どのように生存しているのか。生存の「場」としての環境を,(1)自然生態的環境,(2)社会的環境,(3)象徴的環境に分析上区別して,牧畜民の遊動生活を材料として考えた。
2.ドドスの遊動生活
牧畜民の遊動生活は,植生が貧弱で水の乏しい乾燥地という厳しい自然環境を効率よく利用するための生態学的適応として分析される一方で,人間関係の葛藤や儀礼の開催,隣接集団との「戦い」等の社会的,政治的,宗教的な要因の関与が指摘されてきた。
ドドスはウガンダ北東部のケニア,スーダンとの三国国境地域に住むウシ牧畜民である。生産・消費の単位は父系の拡大家族であり,集落(ere)と家畜キャンプ(awi)に家族のメンバーと保有家畜が分散し,それぞれに自律的に日常生活が営まれている。集落は半定住的であるが,家畜キャンプは数日から1〜2カ月程度の間隔で頻繁に移動を繰り返し,移動距離も数キロメートルから100キロメートルを超える場合もある。したがって,ドドスの遊動生活を形作っているのは実質的には家畜キャンプの移動と言って差し支えない。
3. キャンプ移動の契機
3-1 家畜キャンプの移動の事例
移動の概要をつかむため,2003年6月2日〜7月2日における家畜キャンプ移動の事例を紹介した(表1,河合 2006:185)。
表1 家畜群の移動の経緯(2003.6.2-7.2)
| 移動日 | 移動先(地名) | 移動の「理由」(公式見解) | 滞在日数 |
| ①6月2日 | モルアンガピオン | 預言者の指示 | 1 |
| ②6月3日 | モルエセ | エンゲレイ(含塩土壌)の摂取 | 4 |
| ③6月7日 | モルアンガピオン | 上記,完了 | 7 |
| ④6月14日 | ロクワカラモエ | 腸占いの啓示 | 6 |
| ⑤6月20日 | モルアンガピオン | 前設営地の地形的条件が劣悪なため | 12 |
| ⑥7月2日 | ロイタ | 腸占いの啓示 |
この期間の家畜キャンプの移動は,形の上ではモルアンガピオンを基点に,モルエセとロクワカラモエの2カ所をそれぞれ往復したうえでロイタへ移動しているが,これらの移動の契機となったのは,エンゲレイの摂取が家畜飼育に関与する生態的な理由であることを除けば,預言者の指示や腸占いの啓示を直接的な契機としていた。こうした具体例を踏まえ,移動の契機となったのはどのような環境の変化なのか,ドドスの家畜キャンプ移動が3つの質的に異なる環境との相互作用としてあらわれる様子を追った。
3-2 自然生態的環境
上記の事例にはなかったが,主として草や水の時空間配置の変化に基づくキャンプ移動がある。また,事例のように家畜の成育に不可欠とされている塩分を含む草や土を定期的に採食させるためにキャンプ移動をすることがある。移動の契機となるこれらの要素を含む環境を「自然生態的環境」と呼ぶ。
ドドスにおける自然生態的環境は,降水量の少ない乾燥・半乾燥地における不規則で不安定で予測の難しい気まぐれな気象条件のもとにあり,しばしば旱魃に見舞われる。また,年間通じてほとんど雨の降らない大旱魃に襲われることもある。他方,大雨や洪水も問題となっており(2000年以降に目立つようになった。エルニーニョなど地球規模の気象変動,地球温暖化の影響による現象か?),鉄砲水で家畜が溺死したり,牧草地が長期間水浸しになったり泥水が流れ込むことによって牧草が生えなくなる(根腐れ?)等の被害がある。
自然生態的環境への適応が果たされないと家畜群の維持に支障を来し,ひいては人間の生存が脅かされる。このことを人びとはよく認識しており,自然生態的環境の変化(劣化)への「気づき」により,緊急に対処法が話し合われ,キャンプ移動が決行される。
そもそも,乾燥地という,そのままでは利用できない土地を,家畜を媒介することによって,人間の生息域としてきた乾燥地適応としての牧畜という生業の成立に関わる位置づけは普遍的なものである。だが,「水と草を求めて遊動する牧畜民」というイメージは素朴であり,現実的ではない。自然生態的環境の条件からは移動する必要がないのに移動したり,移動先の条件が自然生態的環境としては厳しすぎるにもかかわらず,あえて困難な場所へ移動したりすることもあるからである。事例では6回の移動の内3回が預言者の指示や腸占いの結果(啓示)がキャンプ移動の契機となっていた。次に述べる社会的環境や象徴的環境への対処を優先する事態が頻繁に起こっているのである。
3-3 社会的環境
ヒトが集団で生きる以上,環境は他者との相互作用の「場」である。そうした他者=同種他個体との相互作用の働く「場」を「社会的環境」と呼ぶ。社会的環境の要素である他者の作用によるキャンプ移動として,自民族内で放牧地を集中させない等の調整は常に意識的にされているが,隣接する他民族集団との関係もキャンプ移動に強く作用する。他集団のレイディングを避けるための移動が実に頻繁なのである。
「(家畜キャンプは)一カ所に長くとどまらない」という言説はよく耳にするが,その理由としては,「草や水が不足する」といった自然生態的環境の諸要素の作用よりもむしろ,「長く滞在すると家畜の居場所や放牧ルートが敵に知られ,放牧中の待ち伏せやキャンプ狙いの襲撃といったレイディングの危険が増すからだ」といった説明の方がより多く聞かれた。キャンプ移動は,自然生態的環境の要素である地勢や植生,採食地や給水地の状態等,移動先が放牧地やキャンプ設営地として適当かどうかの条件を満たしつつ,レイディングへの警戒とも不可分である。レイディングは牧畜民の生業基盤である家畜を群れごと奪われる,人びとの生存に直接的に関わる重大事であり,さまざまな方法でこの危険に対処し,また,キャンプ移動が決行されている。
社会的環境を形作る言説情報や目撃情報とともに,預言者の言葉や腸占いの啓示はレイディングの危険を「知る」よすがとなっていた。それは「兆し」「予兆」「気づき」といったものでもあるが,これらをもとに家畜を守るための行為が選ばれ,実践される。これについて次に述べる。
3-4 象徴的環境
自然生態的環境,社会的環境に加えて,ヒトには「いま,ここ」ではない,時空間を越えて,想像力によって創造される環境がある。これを「象徴的環境」と呼ぶ。ドドスにおいて象徴的環境を作り上げている諸要素の筆頭は「啓示」である。啓示は,主として,(1)預言者(呪医emuron)による夢見やサンダル占い,(2)腸占い,(3)個人的な夢見やサンダル占いなどの結果としてもたらされるが,象徴的環境の要素として日常的であり,かつ人びとが絶大な信頼を置くのは「腸占い」である。キャンプ移動に,腸占いが強くかかわる事例には事欠かない。
「啓示」は,近い将来の出来事(主として危険)を示唆するが,それは現実味を帯びた事態=現実問題として,人びとはさまざまな方法でこれに対処する。家畜の供犠を伴う大規模な儀礼の開催,特定の場所への呪物の設置,まじない的な儀礼的行為等とともに,日帰り放牧のルート変更や家畜キャンプの移動がここに含まれる。象徴的環境の諸要素は,「草がなくなった」「井戸の水が涸れた」などの自然生態的環境の要素の変化と同じ地平の出来事として作用するし,どこそこでレイディングがあったといった言説情報や足跡などの痕跡や人影などの目撃情報によるレイディングの「兆し」といった社会的環境を補強する要素として,人々の活動/行為選択に作用する。象徴的環境の諸要素は,「(生物学的)生存」にとってはしばしば明らかに過剰な行為である。だが,それは,人びとにとっては「生存」に直結する現実的な課題として迫って(作用して)くる事態であり,絶対に何らかの対処をすべき作用である。象徴的環境は想像力によって構築される世界だが,現実の物理空間を「超越」して
3-5 レイヤーとしての環境
以上,3つの環境は,それぞれに独立しているのではないことは明らかである。また,「遊動」という生活様式にとって,どれがより優越しているわけでもない。地理空間上にある特定の場所は,自然生態的環境としてあるとともに,そこは社会的環境が投影された場所でもあり,象徴的環境として意味づけられた場所でもある。ドドスの遊動生活は,そうした質の異なる3つの環境のレイヤー構造=重層的な「場」に展開されるのであり,それぞれの環境からの作用を受けながら,それに対処するように,つまり3つの環境との間の相互作用として現れたものとしてある。
4. 環境の要素としての他者(他民族集団):その存在様態
キャンプ移動の契機となった他者はしばしば「情報」によって生成する存在であった。ここで言う情報は,(1)言説情報や目撃情報および(2)夢見や腸占いによる「啓示」という2種類が区別できる。前者は社会的環境の要素としての他者,後者は象徴的環境の要素としての他者という異なる2つの他者の存在様態を生成する。だが,この2つを人びとが異なる存在様態としてとらえているかどうかは不明である。これら2つが意味をもつのは,他者がレイディングを仕掛けてくる事態に際してであり,これに対処するためにパトロールの強化,儀礼の開催,呪物の設置,放牧コースの変更等をし,最終的には家畜キャンプの移動が選択される。牧畜民の遊動生活は自然生態的環境への適応であるとともに,隣接してともに生きる他者と相互作用する社会的環境や象徴的環境に大きく左右されていたのである。
こうした環境世界のあり方は,ヒトに特異的か?トマセロ(2013:53)によれば,「ヒトは他の類人猿のような物理的・社会的世界を生きているだけでなく,自分たち自身で作りあげた制度的・文化的世界…を生きている」。だが,これまでの共同研究を通じて制度的・文化的世界も萌芽的であれ霊長類にも認められることをわれわれは明らかにしてきた。人間は言葉を持つが故に,言葉が前面に出てしまうが,言葉を使う以前に未来についての「兆し」「予兆」「気づき」は,環境世界の諸要素の作用として既にあるのではないか。そのとき近い未来を「知る」術は霊長類の環境との相互作用と異なるといえるのか。検討の余地がある。
5.おわりに
結論めいたことを言う準備はできなかったが,一連の共同研究を通じて考えようとしてきたのは,隣接する民族集団が「敵対しつつも共存している」状況をいかに理解できるかであり,それは「他者とともに生きる」術(=高度な社会性という能力)の進化史的基盤を問うものであった。本発表で検討してきた「環境」の生成,成立とその利用がそれといかに関わるのかという問いは,今後の課題としたい。
【引用文献】
- 足立薫 2016「環境の他者へ—平衡と共存の行動学試論」『他者—人類社会の進化』京都大学学術出版会,京都
- — ,2013「役割を生きる制度—生態的ニッチと動物の社会」『制度—人類社会の進化』京都大学学術出版会,京都
- M.トマセロ 2015『ヒトはなぜ協力するのか』勁草書房,東京
- 船曳建夫 2016「苦悩としての他者—三者関係と四面体モデル」河合香吏編『他者—人類社会の進化』京都大学学術出版会,京都
- — 2009「人間集団のゼロ水準—集団が消失する水準から探る,関係の意味,場と構造」『集団—人類社会の進化』京都大学学術出版会,京都
- 河合香吏 2006「キャンプ移動と腸占い—ドドスにおける隣接集団との関係をめぐる社会空間の生成機序」西井凉子・田邊繁治編『社会空間の人類学—マテリアリティ・主体・モダニティ』世界思想社,京都
第6回研究会での発表内容
1.「食性の変化と調理加工の共同:食が社会にひらかれるとき」(杉山祐子)
1. はじめに
ヒトの食の特徴は、食物となるものをそれが生育している環境から採/獲ったあと、口に入れるまでの時間がサルと比べて長いことである。この間に獲物の解体や植物性食材の加工処理、調理がおこなわれるが、その過程には、それぞれがとってきた食物を集め、分配する行為を伴っている。調理加工は食物となるものを「食事」に変換し、そこに社会的な意味合いを生み出して「共食」の素地をつくる。
この発表では、料理がヒト化をうながしたとするいわゆる「料理仮説」を視野にいれつつ、火を使う調理そのものだけでなく、調理にいたるまえの加工処理作業を重視して、環境や食性の変化と共同の様態について検討した。
それというのも、民族誌的には農耕民ベンバ社会でも、採集狩猟民サンの社会でも、火を使う調理にいたるまえの加工処理段階で、女性たちの集いが顕著にみられるからである(今村薫 2010、杉山祐子 1987)。とくに穀物を主食とする農耕民では、穀物を脱穀精製する作業やキャッサバなどの毒抜きなどの加工処理作業に対して、農耕そのものにかけるよりも多くの手間と時間を費やしており、その作業が女性たちの集う機会の大半を占めている。ここでは、料理によって食が社会に開かれるという側面が生業形態にかかわらず共通するという前提にたちつつ、定住化や環境への恒常的な働きかけとその変化が、共同の様態とどのように関わりあうのかを考える緒をみいだそうとするものである。
2. 「料理仮説」、人類史における食性の変化
リチャード・ランガム(2009)は、火の使用の起源がこれまで考えられていたより大きく遡り、火による調理がヒト化の大きな要因だったとする「料理仮説」を唱えた。ジェンダーの観点から発生過程についての見解には疑問が大きいものの、料理と性別分業の発生についての指摘も重要である。
料理仮説を考古学的証拠から検討した鵜澤和宏(2012)によれば、料理はヒトの食性の幅を広げ、寒冷地を含む多様な環境へのヒトの拡散も可能にしたはずだという。11000年前にコムギ栽培が始まった西アジアを起点に、穀物を中心とした食性が広がったが、それは「人類は料理によって作られるという状況が現実のもの(鵜澤2012:60)」となったできごとだった。
火による調理が一般化したヒトの食は、農耕の開始以前から、分業を含む他者への依存を個体としての生存の前提とするという意味でヒト的な社会性をまとったといえるが、人類史のなかではきわめて新しい穀物食への変化はそれをさらに大きく展開させ、共同性や分配のしくみの複雑化、さらには階層化や権力の出現にも深く関わる。北アメリカ先住民や日本の縄文社会にみられる堅果類等の利用もまた、技術や道具の精緻化と専門化、社会組織の複雑化や重層化を促したことが知られている。
3. 極相化する環境と変化
一時期に大量の収穫がある穀物や堅果類を食の中心にすえるようになることは、特定の場所にヒトが恒常的な働きかけを続け、獲得すべき資源が持続的に得られるような環境を実現していくことと不可分である。当該の資源を効率よく安定して得るためのさまざまな働きかけを通して、環境とヒトの活動との相互依存が促される。その結果、当該の資源の生産・再生産にもっとも適し、そのしくみを組み込んだ極相的な環境が生み出される。それは、西田正規(1996)が述べたように、定住生活の出現と切り離せない。
西田は小規模な定住集落を生み出していた縄文社会にすでに、ブナ科の澱粉質種子類を産する植物の栽培化が始まっていたことを指摘したが、定住集落の周辺環境の二次林化や集落周辺土壌の豊栄養化が栽培化の素地となり、栽培化−人間活動と植物との共生関係が成立したと述べる。澱粉質種子類を食の中心にし、その利用に適した環境の再生サイクルを安定的に維持することによって、縄文中期には大集落が構成され、集落周辺の二次林を含む多様な環境を巧みに利用して、バラエティ豊かな食を安定的に得ていたことが知られている。道具・技術の精緻化やある種の専業化、集落の立地や儀礼などが同調して精緻に発達した背景には、こうした人間活動と植物との共生関係をとおした環境の極相化があるとかんがえたい。しかし、この極相的環境利用のしくみは縄文晩期におこった急激な気候の冷涼化による環境変動に対応できずに衰退し、東日本での集落規模の縮小や西日本での人口減少につながったという(上條2016)。
4.現代アフリカ農村における調理加工の共同と作物の変化
4-1.食材と調理加工の共同
一定期間の定住を基盤とする栽培化−農耕は、狩猟採集と比べると、農耕は食料資源を得たあとで、より多くの手間と労力を費やすことを前提に持続可能性を保つ資源利用の方法である。なかでも穀物を中心とする食性は、調理加工の共同の必要性をより高めるが、必要性の大きさや共同の様態は作物によっても異なる。現代のアフリカ農村の事例を見ても、有毒キャッサバなどを除くイモ類やバナナなどは、随時収穫でき、加工処理がほとんど必要ない作物である。これらは共同での作業もおこなわれるが、個別で手軽に調理ができる特徴をもつ。
これに対して、とれる時期が決まっており、一斉に収穫しなければならない穀物類は、共同で作業にあたる必要性が高く、その加工処理には長時間を要する。とくにいくつもの道具と手間を必要とする穀物の脱穀や精選、製粉などの作業では、少量を個別に処理するより、ある程度の量をまとめて作業するほうがより効率的である。同じ穀物でも、粒の大きさや脱粒の手間によってもその必要性は異なる。作業効率を考えて機能的にみれば、穀物の加工処理は、作業を共同する必要性がより高いといえる。
効率とは別に、作業に従事する側にとっても、単調で長く続く作業は、ほかの人と共同でするか、作業を共同しないまでも同じ場所に集まって従事することで、はるかに楽で楽しいものになる。それはベンバの村の女性が「ひとりで黙ってこんなに長い時間(穀物を)搗き続けるなんて、とても毎日はがまんできないわよ」と言ったように、この作業をただのつらい仕事におとしめないことでもある。
それはまた、加工作業に続く調理と食事に、自然に参加できる状況をも整えてくれる。
ただ、実際にそこに残って食事を共にするおとなは、当該の世帯の近い母系親族やごく親しい友人で、ほかのおとな女性は食事の始まる前に何気なくその場を去るか、勧められても「ありがとう」と断ることが多い。
当該の世帯との関係にかかわらず、食事の供与にあずかるのは5、6歳くらいまでの子どもたちである。子どもたちは加工作業に一緒に来た母親たちがその場からいなくなっても、あたりまえのように食事に参加し、満足するまで食べていく。こうした観察例からかんがえると、調理加工の共同とそれに続く共食は、子どもたちが十分な食事を得る機会を保証するという意味でも重要だといえる。
他方、別稿で明らかにしたように、ベンバの村でおとな女性が調理加工の場に集うことをやめるのは、共に暮らしつづける気持ちがなくなり村を離れるつもりだという意思表明にほかならないことを考え合わせると(杉山1987)、おとな女性たちにとっての調理加工の共同は、集まる、場を共有するということ自体にも重要な意味があることを確認しておきたい。
4-2.共同の様態の変化と生計
オードリーリチャーズは、1930年代の北ローデシア(現ザンビア)のベンバ社会における共同調理(joint cooking)グループについて記している(Richards, 1939:132)。共同調理は女性の労働を大きく軽減し、農繁期には畑からオカズを持ち帰る女性たちと主食の準備をする女性たちにわかれて調理加工を分担・共同するようすがみられたようだ。リチャーズはこうした共同調理がそれぞれの世帯の生計の安定に大きく寄与していることや、作業をともにすることによる知識の伝達などの機能にも注目した。しかし、現金経済の浸透による生活の変化によって共同調理グループの規模が縮小し、調理加工が個別化したため、こうした利点が享受できなくなり、食料不足のときに困窮する世帯が生じていることを報告している。
リチャーズの記述と類似の事例が1980年代後半〜1990年代のベンバ社会でも観察できた。このころ、ハイブリッド種トウモロコシ栽培が急速に広がったが、その受容は製粉機械の導入と同時におきた。脱粒から製粉作業までほとんどを個別におこなうので、女性たちが集まる機会が少なくなった。その影響は、トウモロコシ栽培に力を入れ始め、家族内での女性の活動が増えた世帯に大きく現れた。調理加工作業で他の女性たちと集うことが少なくなった世帯には、作業中にちょっとした食材のやりとりをしたり、それに続く共食にあずかる機会が減ったのである。トウモロコシ栽培に移行中の世帯では食物のバラエティが狭まっていることを示す報告もなされている(Bolt R., Silavwe M., and Bolt 1987)。
上述のとおり、調理加工の共同は、作業効率だけでなく集まることじたいに重要な意味をもつのだが、それはこうした例からもわかるように、女性のあいだでの食料や仕事の分担などといった機会を提供して、生計の安定や食物バラエティの確保につながるという機能的な重要性ももっている。同じ穀物食であっても、作物とその加工処理方法が変わると共同の様態も変化し、それが個々の食料獲得にも大きな影響を与えるといえる。
5.おわりに
ヒトが食物となる資源を「とる行動」とそれを食物として「食べる(口に入れる)行動」は、あいだに調理加工のプロセスが埋め込まれたことによって、それぞれ別の社会的位相におかれることになった。調理加工は資源の加工プロセスであると同時に、個体の生存維持の要である食に他者が介在するプロセスでもある。
穀物食への食性の変化は、調理加工の手間と時間をより長期化させ、共同の必要性を高めた。ここでの共同はそれぞれのメンバーがとってきたものを集め、食事という文化的生産物として統合する役目を担うだけでなく、なにげない分かち合いや共食につながる場を形成する。またひとが集まる場であること自体にも社会的に重要な意味合いが与えられる。この共同の様態は、当該の食材の特徴や加工処理方法にも応じて変化しやすく、共同の様態の変化が食材となる作物の選択や分かち合いの様態の変化とも深く関わっている。
ひるがえって、高度に近代化・産業化が進んだ現在の日本の食のありようをみてみると、そこでは加工処理作業の産業化と調理の家族化、個別化が顕著にあらわれている。「とる行動」と「食べる行動」が別の社会的位相にあり、食べるために料理が不可欠というヒト的な食の状況は変わらないが、近代化のなかで加工調理の大部分が産業化されたために、食べる前に共同での加工処理作業を必要とするものはほぼなく、すぐに調理できたりすぐに食べられたりするものに置き換わって、「食べる行動」をめぐる諸々が家庭のなかに閉じ込められた。現代日本の食は、社会に開かれたものから、世帯に閉じたもの、あるいはきわめて個別化したものに変質しており、動物としての生存環境としてはかなり極限的な状況にあるとも考えられる。こうした現状にあって、食をふたたび社会に開こうとするいくつもの試みがなされている事例もあり、これらもまた、人類史をかんがえるうえで、興味深い点である。
引用文献
ヒトの食の特徴は、食物となるものをそれが生育している環境から採/獲ったあと、口に入れるまでの時間がサルと比べて長いことである。この間に獲物の解体や植物性食材の加工処理、調理がおこなわれるが、その過程には、それぞれがとってきた食物を集め、分配する行為を伴っている。調理加工は食物となるものを「食事」に変換し、そこに社会的な意味合いを生み出して「共食」の素地をつくる。
この発表では、料理がヒト化をうながしたとするいわゆる「料理仮説」を視野にいれつつ、火を使う調理そのものだけでなく、調理にいたるまえの加工処理作業を重視して、環境や食性の変化と共同の様態について検討した。
それというのも、民族誌的には農耕民ベンバ社会でも、採集狩猟民サンの社会でも、火を使う調理にいたるまえの加工処理段階で、女性たちの集いが顕著にみられるからである(今村薫 2010、杉山祐子 1987)。とくに穀物を主食とする農耕民では、穀物を脱穀精製する作業やキャッサバなどの毒抜きなどの加工処理作業に対して、農耕そのものにかけるよりも多くの手間と時間を費やしており、その作業が女性たちの集う機会の大半を占めている。ここでは、料理によって食が社会に開かれるという側面が生業形態にかかわらず共通するという前提にたちつつ、定住化や環境への恒常的な働きかけとその変化が、共同の様態とどのように関わりあうのかを考える緒をみいだそうとするものである。
2. 「料理仮説」、人類史における食性の変化
リチャード・ランガム(2009)は、火の使用の起源がこれまで考えられていたより大きく遡り、火による調理がヒト化の大きな要因だったとする「料理仮説」を唱えた。ジェンダーの観点から発生過程についての見解には疑問が大きいものの、料理と性別分業の発生についての指摘も重要である。
料理仮説を考古学的証拠から検討した鵜澤和宏(2012)によれば、料理はヒトの食性の幅を広げ、寒冷地を含む多様な環境へのヒトの拡散も可能にしたはずだという。11000年前にコムギ栽培が始まった西アジアを起点に、穀物を中心とした食性が広がったが、それは「人類は料理によって作られるという状況が現実のもの(鵜澤2012:60)」となったできごとだった。
火による調理が一般化したヒトの食は、農耕の開始以前から、分業を含む他者への依存を個体としての生存の前提とするという意味でヒト的な社会性をまとったといえるが、人類史のなかではきわめて新しい穀物食への変化はそれをさらに大きく展開させ、共同性や分配のしくみの複雑化、さらには階層化や権力の出現にも深く関わる。北アメリカ先住民や日本の縄文社会にみられる堅果類等の利用もまた、技術や道具の精緻化と専門化、社会組織の複雑化や重層化を促したことが知られている。
3. 極相化する環境と変化
一時期に大量の収穫がある穀物や堅果類を食の中心にすえるようになることは、特定の場所にヒトが恒常的な働きかけを続け、獲得すべき資源が持続的に得られるような環境を実現していくことと不可分である。当該の資源を効率よく安定して得るためのさまざまな働きかけを通して、環境とヒトの活動との相互依存が促される。その結果、当該の資源の生産・再生産にもっとも適し、そのしくみを組み込んだ極相的な環境が生み出される。それは、西田正規(1996)が述べたように、定住生活の出現と切り離せない。
西田は小規模な定住集落を生み出していた縄文社会にすでに、ブナ科の澱粉質種子類を産する植物の栽培化が始まっていたことを指摘したが、定住集落の周辺環境の二次林化や集落周辺土壌の豊栄養化が栽培化の素地となり、栽培化−人間活動と植物との共生関係が成立したと述べる。澱粉質種子類を食の中心にし、その利用に適した環境の再生サイクルを安定的に維持することによって、縄文中期には大集落が構成され、集落周辺の二次林を含む多様な環境を巧みに利用して、バラエティ豊かな食を安定的に得ていたことが知られている。道具・技術の精緻化やある種の専業化、集落の立地や儀礼などが同調して精緻に発達した背景には、こうした人間活動と植物との共生関係をとおした環境の極相化があるとかんがえたい。しかし、この極相的環境利用のしくみは縄文晩期におこった急激な気候の冷涼化による環境変動に対応できずに衰退し、東日本での集落規模の縮小や西日本での人口減少につながったという(上條2016)。
4.現代アフリカ農村における調理加工の共同と作物の変化
4-1.食材と調理加工の共同
一定期間の定住を基盤とする栽培化−農耕は、狩猟採集と比べると、農耕は食料資源を得たあとで、より多くの手間と労力を費やすことを前提に持続可能性を保つ資源利用の方法である。なかでも穀物を中心とする食性は、調理加工の共同の必要性をより高めるが、必要性の大きさや共同の様態は作物によっても異なる。現代のアフリカ農村の事例を見ても、有毒キャッサバなどを除くイモ類やバナナなどは、随時収穫でき、加工処理がほとんど必要ない作物である。これらは共同での作業もおこなわれるが、個別で手軽に調理ができる特徴をもつ。
これに対して、とれる時期が決まっており、一斉に収穫しなければならない穀物類は、共同で作業にあたる必要性が高く、その加工処理には長時間を要する。とくにいくつもの道具と手間を必要とする穀物の脱穀や精選、製粉などの作業では、少量を個別に処理するより、ある程度の量をまとめて作業するほうがより効率的である。同じ穀物でも、粒の大きさや脱粒の手間によってもその必要性は異なる。作業効率を考えて機能的にみれば、穀物の加工処理は、作業を共同する必要性がより高いといえる。
効率とは別に、作業に従事する側にとっても、単調で長く続く作業は、ほかの人と共同でするか、作業を共同しないまでも同じ場所に集まって従事することで、はるかに楽で楽しいものになる。それはベンバの村の女性が「ひとりで黙ってこんなに長い時間(穀物を)搗き続けるなんて、とても毎日はがまんできないわよ」と言ったように、この作業をただのつらい仕事におとしめないことでもある。
それはまた、加工作業に続く調理と食事に、自然に参加できる状況をも整えてくれる。
ただ、実際にそこに残って食事を共にするおとなは、当該の世帯の近い母系親族やごく親しい友人で、ほかのおとな女性は食事の始まる前に何気なくその場を去るか、勧められても「ありがとう」と断ることが多い。
当該の世帯との関係にかかわらず、食事の供与にあずかるのは5、6歳くらいまでの子どもたちである。子どもたちは加工作業に一緒に来た母親たちがその場からいなくなっても、あたりまえのように食事に参加し、満足するまで食べていく。こうした観察例からかんがえると、調理加工の共同とそれに続く共食は、子どもたちが十分な食事を得る機会を保証するという意味でも重要だといえる。
他方、別稿で明らかにしたように、ベンバの村でおとな女性が調理加工の場に集うことをやめるのは、共に暮らしつづける気持ちがなくなり村を離れるつもりだという意思表明にほかならないことを考え合わせると(杉山1987)、おとな女性たちにとっての調理加工の共同は、集まる、場を共有するということ自体にも重要な意味があることを確認しておきたい。
4-2.共同の様態の変化と生計
オードリーリチャーズは、1930年代の北ローデシア(現ザンビア)のベンバ社会における共同調理(joint cooking)グループについて記している(Richards, 1939:132)。共同調理は女性の労働を大きく軽減し、農繁期には畑からオカズを持ち帰る女性たちと主食の準備をする女性たちにわかれて調理加工を分担・共同するようすがみられたようだ。リチャーズはこうした共同調理がそれぞれの世帯の生計の安定に大きく寄与していることや、作業をともにすることによる知識の伝達などの機能にも注目した。しかし、現金経済の浸透による生活の変化によって共同調理グループの規模が縮小し、調理加工が個別化したため、こうした利点が享受できなくなり、食料不足のときに困窮する世帯が生じていることを報告している。
リチャーズの記述と類似の事例が1980年代後半〜1990年代のベンバ社会でも観察できた。このころ、ハイブリッド種トウモロコシ栽培が急速に広がったが、その受容は製粉機械の導入と同時におきた。脱粒から製粉作業までほとんどを個別におこなうので、女性たちが集まる機会が少なくなった。その影響は、トウモロコシ栽培に力を入れ始め、家族内での女性の活動が増えた世帯に大きく現れた。調理加工作業で他の女性たちと集うことが少なくなった世帯には、作業中にちょっとした食材のやりとりをしたり、それに続く共食にあずかる機会が減ったのである。トウモロコシ栽培に移行中の世帯では食物のバラエティが狭まっていることを示す報告もなされている(Bolt R., Silavwe M., and Bolt 1987)。
上述のとおり、調理加工の共同は、作業効率だけでなく集まることじたいに重要な意味をもつのだが、それはこうした例からもわかるように、女性のあいだでの食料や仕事の分担などといった機会を提供して、生計の安定や食物バラエティの確保につながるという機能的な重要性ももっている。同じ穀物食であっても、作物とその加工処理方法が変わると共同の様態も変化し、それが個々の食料獲得にも大きな影響を与えるといえる。
5.おわりに
ヒトが食物となる資源を「とる行動」とそれを食物として「食べる(口に入れる)行動」は、あいだに調理加工のプロセスが埋め込まれたことによって、それぞれ別の社会的位相におかれることになった。調理加工は資源の加工プロセスであると同時に、個体の生存維持の要である食に他者が介在するプロセスでもある。
穀物食への食性の変化は、調理加工の手間と時間をより長期化させ、共同の必要性を高めた。ここでの共同はそれぞれのメンバーがとってきたものを集め、食事という文化的生産物として統合する役目を担うだけでなく、なにげない分かち合いや共食につながる場を形成する。またひとが集まる場であること自体にも社会的に重要な意味合いが与えられる。この共同の様態は、当該の食材の特徴や加工処理方法にも応じて変化しやすく、共同の様態の変化が食材となる作物の選択や分かち合いの様態の変化とも深く関わっている。
ひるがえって、高度に近代化・産業化が進んだ現在の日本の食のありようをみてみると、そこでは加工処理作業の産業化と調理の家族化、個別化が顕著にあらわれている。「とる行動」と「食べる行動」が別の社会的位相にあり、食べるために料理が不可欠というヒト的な食の状況は変わらないが、近代化のなかで加工調理の大部分が産業化されたために、食べる前に共同での加工処理作業を必要とするものはほぼなく、すぐに調理できたりすぐに食べられたりするものに置き換わって、「食べる行動」をめぐる諸々が家庭のなかに閉じ込められた。現代日本の食は、社会に開かれたものから、世帯に閉じたもの、あるいはきわめて個別化したものに変質しており、動物としての生存環境としてはかなり極限的な状況にあるとも考えられる。こうした現状にあって、食をふたたび社会に開こうとするいくつもの試みがなされている事例もあり、これらもまた、人類史をかんがえるうえで、興味深い点である。
引用文献
- Bolt R., Silavwe M., and Bolt M. 1987 Food availability and consumption patterns in the Northern Region of Zambia ARPT trial areas: implications for research extension and policy. ARPT
- Richards, A.I. 1939 Land, Labour and Diet in Northern Rhodesia, International African Institute
- Zeder, M.A. 2011 The Origins of Agriculture in the Near East, Current Anthropology, Vol.52 No.S4:S221-S235
- 今村薫 2010 『砂漠に生きる女たち』どうぶつ社
- 鵜澤和宏 2012 「人類の進化と火」朝倉敏夫編『火と食』ドメス出版 44-66
- 上條信彦 2016 『縄文時代における脱穀・粉砕技術の研究』六一書房
- 杉山祐子 1987 「臼を貸してください」『アフリカ研究』
- 西田正規 1997 「栽培と農耕、出現過程の生態学」『霊長類研究』113:173-181
- ランガム、リチャード 2009 『火の賜物─人は料理で進化した』NTT出版
2.「社会の極限としての孤独性:野生チンパンジーが孤独になるとき」(西江仁徳)
■はじめに
●社会と孤独
これまで集団の集合性を軸に議論されてきた社会構造論から、離散性を軸にした社会性の解離の議論への展開をはかるべく、社会における単独性(孤独)に焦点をあてる。ここで「単独」ではなくあえて「孤独」ということの意味は、単に個体としてあるという意味での「単独性」ではなく、社会の一体性の解体・切断という意味での「解離」(dissociation)や「脱組織化」(disorganization)を、凝集/集合状態のネガではなく生物の社会性の基本的な一側面として主題化したいという目論見にもとづいたものである。
ある先験的な社会的・生態的環境に偶発的に生まれ落ちた生物個体として、社会の一体性の「解離」や「脱組織化」として現れる「孤独」は、その生存を左右する極限的な環境であると考えられる。こうした社会における「孤独」に関する議論は、すでに『集団—人類社会の進化』(河合, 2009)のいくつかの章で(とくに集団の「非構造」の問題として)論じられたテーマではあるが、野生チンパンジーの単独性(孤独性)の現象をいくつかの事例を通して論じ直すことで、社会性と孤独との(理論的)関係についての考察を深めたい。
●構造と非構造 (伊谷, 1993; 足立, 2009)
伊谷 (1987)の社会構造論における「構造」とは、集団の性構成と移籍パターンによって規定された、それぞれの種に固有のBSU(基本的単位集団)のかたちをとって現れる。一方で伊谷 (1993)が晩年に構想した「非構造」は、「構造」におさまりきらない存在者として定義され、BSUの周辺に存在する移籍個体やゴリラのオス集団、鳥の大群や複数の種によって構成される混群などがその例としてあげられてきた。とくに伊谷 (1993)が非構造の典型例として注目したニホンザルのヒトリオスについて、内堀 (2009)はそれが構造によって規定された移籍する性の個体に限られる現象であることから「構造化されたプロセスの一環」であるとしたが、伊谷非構造論を発展的に継承・展開した足立 (2009)は「非構造の孤独」と位置づけた。これは、ニホンザルのヒトリオスが「移籍する性」の個体であり、「移籍する性」は構造によって規定されているために、それが「構造」(内堀, 2009)であるともいえる一方、移籍個体は構造の外の存在者であるために「非構造」(足立, 2009)でもある、という錯綜した位置づけになっていると考えられる。本発表では、父系の社会構造(メスが集団間を移籍する)をもつ野生チンパンジー社会においてこれまで散発的に観察されてきた「オスの集団からの長期離脱にともなう単独生活」に焦点をあて、その非構造性、社会の解体や脱組織化の契機としての働きを「可能態(潜勢態)としての孤独」として描く。実質的に移籍の可能性がないチンパンジーのオスにとって、集団からの離脱にともなう単独生活は「集団の外に出ていくわけにも行かず、かといって簡単に集団に戻ることも難しい」という意味で「出口なし」の極限状態と考えられるが、その状態をチンパンジーのオスたちがどのように過ごしているのかについて過去の報告を参照しつつ検討する。
●チンパンジーの「離合集散」概念の整理
野生チンパンジーは父系の複雄複雌の単位集団を形成する。このチンパンジーの単位集団は、ある程度の期間にわたってメンバーシップの安定した集団であり、メスは性成熟にともなって出自集団から移出し、別の単位集団に移入することが多いが、オスは基本的に移籍しない。
チンパンジーの単位集団は、そのメンバーが常に凝集性を高く保つのではなく、頻繁に「離合集散」する特徴をもっている。(Itoh and Nishida, 2007; Nishida, 1968)。このチンパンジー社会の離合集散性について、黒田 (2009)は「離合集散とは、生態的・社会的に集団から自己を分離できる者たちがつくる集団現象」であり、「単独になる力でもあるという意味で、「単独性」といってもよい」と述べている (黒田, 2009)。本発表で検討する「チンパンジーのオスの長期的離脱と単独生活」についても、こうしたチンパンジー社会の離合集散性の連続性のなかに位置づける。離合集散には一般に「空間スケール」によって記述できる側面と、「時間スケール」によって記述できる側面があり、この二つのスケールを区別することで離合集散の一つの極限的状態としての「孤独」を位置づけることが可能になる。離合集散の空間スケールとしては、視界内に含まれる個体の集まりである「対面パーティ」と、視界外の個体を含むゆるやかなまとまりとしての「遊動パーティ」が区別される (Itoh and Nishida, 2007)。「対面パーティ」の構成やサイズは、目まぐるしく変化するときとそうでないときがある。「遊動パーティ」は互いに声が聴こえる範囲にいておおまかに遊動の方向が一致したまとまりであり、遊動パーティのサイズはチンパンジーの主要な食物資源である果実の密度に依存して季節的に大きく変動することが知られている (Itoh and Nishida, 2007)。離合集散の時間スケールとしては、まず「1日〜数日の中での離合集散」のフェーズがあり、ここでは主に対面パーティのサイズや構成の変化として離合集散が現れる。次に、「季節的な離合集散」は数カ月程度の単位で起こり、主に遊動パーティのサイズや構成の変化として離合集散が現れる(集合季/分散季)。最後に、「集団からの長期的な離脱やその後の復帰」は数カ月〜数年の単位で起こる現象であり、特定の個体の集団からの長期的な離脱として離合集散が現れる。このように、日常的な「対面パーティ/遊動パーティの構成やサイズの変化」を時間的・空間的に引き伸ばしていったところに、本発表で扱うような「特定の個体の長期的な離脱にともなう単独生活(およびその後の復帰)」という現象を位置づけることができるだろう。
●チンパンジーのオスの単独性
チンパンジー社会は複雄複雌で父系の単位集団をもち、メスが集団間移籍をするという「構造」をもつが、この構造については社会生態学や霊長類社会学から説明がなされてきた。
社会生態学は、チンパンジー社会の「オスの集合性」と「メスの単独性」を対置する (Wrangham, 1987)。メスにとっての最重要資源である食物資源(果実)の分布様式への適応として、メスの単独性と狭いコアエリアの利用が説明され、そうしたメスが食物資源防衛と子殺しリスク低減のためにオスを集団内に囲い込む、とされる。一方オスは、オスにとっての最重要資源である繁殖資源(メス)の分布様式への適応として、オス同士が高い集合性を維持し、隣接他集団からメスを共同で防衛する、と説明される。この社会生態学によるチンパンジーの社会構造の説明では、オスの単独性という現象自体が問題にされていない。
霊長類社会学においても同様に、チンパンジー社会におけるオスの集合性に焦点をあてた議論が展開されている (黒田, 2009)。黒田 (2009)は、チンパンジーのオスは「優位志向者」であるとし、自らの順位を高めるためには他のオスとの連合が重要であり、またその連合関係の維持に常に気を配り確かめる必要からオス同士は高い集合性を維持する必要がある、と論じた。そのため、「チンパンジーのオスたちは孤立を恐れ、敵愾心を抑制し、関係確認の交渉を繰り返す」こと、また本発表との関連では、「チンパンジーのオスが順位争いで負けたあと単独行動するのは「敗者の自由」に近い」とまで述べている (黒田, 2009)。つまり、優位志向者であるチンパンジーのオスには「集団化の必然性」があり、「優位」が成立する条件としてチンパンジーのオスは集まらなければならず、優位であるメリットの対象であるメスを集めなければならない、というのである (黒田, 2009)。
しかし、これらはチンパンジーのオスの生活史をかなり単純化して捉えているのではないだろうか?例えば、タンザニア・ゴンベのチンパンジーの社交性の性差について、グドール (1986)は「オスの単独性」と「メスの非単独性(高い社交性)」について論じている。また同じくグドール (1986)はオスの優位志向性についても、すべてのオスが自らの社会的地位の上昇を目指すわけではなく、ごく短期間しか努力しないオスやすぐに順位上昇をあきらめてしまうオス、さらには自分の社会的地位にまったく関心を払っていないようにみえるオスについても言及し、「チンパンジーのオス=優位志向者」という単純な図式におさまらないオスの多様なあり方について論じている。
そこで、本発表では、タンザニア・マハレでこれまで観察されたオスのチンパンジーの単独生活の事例を検討することを通して、「移籍の可能性がなく」「高い集合性をもつ」とされるチンパンジーのオスの行動様式に「可能態(潜勢態)としての孤独」が含まれていることを示し、さらにその「可能態としての孤独」が社会にもたらすものについて議論する。
■タンザニア・マハレのチンパンジーのオスの単独行動事例
●集団からの長期的離脱→単独行動の例
・アルファオス交代にともなう集団からの長期的離脱の事例は、マハレではK集団で1例 (Nishida, 1983; 西田, 1981)とM集団で3例 (Uehara et al., 1994a; 上原, 1994; 保坂 and 西田, 2002; 西江, 2013, 2016)、タンザニアのゴンベ (Goodall, 1986, 1992)でも詳細に観察されているが、それほど頻繁に起こるわけではない。また、アルファオス以外の個体の長期的離脱・単独行動の事例もマハレでは2例観察されている (Nishida et al., 1995; Uehara et al., 1994b, 1994a, 上原, 1994, 2003)。また、マハレとゴンベ以外ではオスの長期的離脱や単独行動についての報告はない。
■考察
●チンパンジーのオスの単独生活を「可能態としての孤独」として見る
移籍の可能性が実質的にないと考えられるチンパンジーのオトナオスの単独生活は、ある意味では「出口なし」の極限的状態と考えられる。その最たる例が、K集団消滅後も長期にわたって単独での生活を余儀なくされたリモンゴの例といえるだろう。また、カジュギのように集団に復帰することなく消失した例もあり、チンパンジーのオスにとって単独生活から集団への復帰は容易ではないことが推察される。
ところがそうした「極限的状態」であると考えられるにも関わらず、実際に観察された単独生活中のオスのチンパンジーの様子は意外にも「普通」であり、おそらく際立った変化があまり見られないため過去の報告にも記載が少ない。単独生活中のカソンタを観察した西田 (1981)の記述や、単独生活中のカジュギの様子の記述 (上原, 1994; 西田, 1981)、また私自身が単独生活中のファナナを観察した印象としても (西江, 2016)、単独生活を送っているときのオスたちの姿は、離脱前と比べてもほとんど変化がなく、健康に過ごしている様子が報告されている。
このように、チンパンジーのオスは単独生活中でもそれ以前ととくに変わりなく「普通に」生活することが可能で、しかもそうした単独生活を数カ月から数年にわたって継続することすら可能である。チンパンジーのオスは「いつもそうする」わけではないが、「必要なときには集団から離脱して単独に近い生活を長期にわたって維持できる」という意味で、「潜勢態としての能力ないし性向」(内堀, 2009)としての孤独を、その行動様式のバリエーションのなかに備えているということができるだろう。こうした「可能態(潜勢態)としての孤独」をチンパンジーのオスの行動様式として描くことは、集団への「復帰」や優位志向性を前提としたり単独生活を「敗者の自由」(黒田, 2009)として描いたりするようなこれまでのチンパンジーのオスのステレオタイプとは異なる、新たな「チンパンジーのオスの可能態(潜勢態)としての社会性」を指し示しているといえるだろう。この「可能態(潜勢態)としての孤独」が社会にもたらしうるものの広がりを検討することで、社会構造の変化や単位集団の分裂・再編成の可能性、社会構造の進化史的祖型からの単独を含む小集団の析出の可能性、複数のコミュニティへの個体の帰属の可能性など、社会の柔軟性や新たな構造/非構造への移行といった「集団性の解体・再編成」の契機を探ることができると考えられる。
■文献
●社会と孤独
これまで集団の集合性を軸に議論されてきた社会構造論から、離散性を軸にした社会性の解離の議論への展開をはかるべく、社会における単独性(孤独)に焦点をあてる。ここで「単独」ではなくあえて「孤独」ということの意味は、単に個体としてあるという意味での「単独性」ではなく、社会の一体性の解体・切断という意味での「解離」(dissociation)や「脱組織化」(disorganization)を、凝集/集合状態のネガではなく生物の社会性の基本的な一側面として主題化したいという目論見にもとづいたものである。
ある先験的な社会的・生態的環境に偶発的に生まれ落ちた生物個体として、社会の一体性の「解離」や「脱組織化」として現れる「孤独」は、その生存を左右する極限的な環境であると考えられる。こうした社会における「孤独」に関する議論は、すでに『集団—人類社会の進化』(河合, 2009)のいくつかの章で(とくに集団の「非構造」の問題として)論じられたテーマではあるが、野生チンパンジーの単独性(孤独性)の現象をいくつかの事例を通して論じ直すことで、社会性と孤独との(理論的)関係についての考察を深めたい。
●構造と非構造 (伊谷, 1993; 足立, 2009)
伊谷 (1987)の社会構造論における「構造」とは、集団の性構成と移籍パターンによって規定された、それぞれの種に固有のBSU(基本的単位集団)のかたちをとって現れる。一方で伊谷 (1993)が晩年に構想した「非構造」は、「構造」におさまりきらない存在者として定義され、BSUの周辺に存在する移籍個体やゴリラのオス集団、鳥の大群や複数の種によって構成される混群などがその例としてあげられてきた。とくに伊谷 (1993)が非構造の典型例として注目したニホンザルのヒトリオスについて、内堀 (2009)はそれが構造によって規定された移籍する性の個体に限られる現象であることから「構造化されたプロセスの一環」であるとしたが、伊谷非構造論を発展的に継承・展開した足立 (2009)は「非構造の孤独」と位置づけた。これは、ニホンザルのヒトリオスが「移籍する性」の個体であり、「移籍する性」は構造によって規定されているために、それが「構造」(内堀, 2009)であるともいえる一方、移籍個体は構造の外の存在者であるために「非構造」(足立, 2009)でもある、という錯綜した位置づけになっていると考えられる。本発表では、父系の社会構造(メスが集団間を移籍する)をもつ野生チンパンジー社会においてこれまで散発的に観察されてきた「オスの集団からの長期離脱にともなう単独生活」に焦点をあて、その非構造性、社会の解体や脱組織化の契機としての働きを「可能態(潜勢態)としての孤独」として描く。実質的に移籍の可能性がないチンパンジーのオスにとって、集団からの離脱にともなう単独生活は「集団の外に出ていくわけにも行かず、かといって簡単に集団に戻ることも難しい」という意味で「出口なし」の極限状態と考えられるが、その状態をチンパンジーのオスたちがどのように過ごしているのかについて過去の報告を参照しつつ検討する。
●チンパンジーの「離合集散」概念の整理
野生チンパンジーは父系の複雄複雌の単位集団を形成する。このチンパンジーの単位集団は、ある程度の期間にわたってメンバーシップの安定した集団であり、メスは性成熟にともなって出自集団から移出し、別の単位集団に移入することが多いが、オスは基本的に移籍しない。
チンパンジーの単位集団は、そのメンバーが常に凝集性を高く保つのではなく、頻繁に「離合集散」する特徴をもっている。(Itoh and Nishida, 2007; Nishida, 1968)。このチンパンジー社会の離合集散性について、黒田 (2009)は「離合集散とは、生態的・社会的に集団から自己を分離できる者たちがつくる集団現象」であり、「単独になる力でもあるという意味で、「単独性」といってもよい」と述べている (黒田, 2009)。本発表で検討する「チンパンジーのオスの長期的離脱と単独生活」についても、こうしたチンパンジー社会の離合集散性の連続性のなかに位置づける。離合集散には一般に「空間スケール」によって記述できる側面と、「時間スケール」によって記述できる側面があり、この二つのスケールを区別することで離合集散の一つの極限的状態としての「孤独」を位置づけることが可能になる。離合集散の空間スケールとしては、視界内に含まれる個体の集まりである「対面パーティ」と、視界外の個体を含むゆるやかなまとまりとしての「遊動パーティ」が区別される (Itoh and Nishida, 2007)。「対面パーティ」の構成やサイズは、目まぐるしく変化するときとそうでないときがある。「遊動パーティ」は互いに声が聴こえる範囲にいておおまかに遊動の方向が一致したまとまりであり、遊動パーティのサイズはチンパンジーの主要な食物資源である果実の密度に依存して季節的に大きく変動することが知られている (Itoh and Nishida, 2007)。離合集散の時間スケールとしては、まず「1日〜数日の中での離合集散」のフェーズがあり、ここでは主に対面パーティのサイズや構成の変化として離合集散が現れる。次に、「季節的な離合集散」は数カ月程度の単位で起こり、主に遊動パーティのサイズや構成の変化として離合集散が現れる(集合季/分散季)。最後に、「集団からの長期的な離脱やその後の復帰」は数カ月〜数年の単位で起こる現象であり、特定の個体の集団からの長期的な離脱として離合集散が現れる。このように、日常的な「対面パーティ/遊動パーティの構成やサイズの変化」を時間的・空間的に引き伸ばしていったところに、本発表で扱うような「特定の個体の長期的な離脱にともなう単独生活(およびその後の復帰)」という現象を位置づけることができるだろう。
●チンパンジーのオスの単独性
チンパンジー社会は複雄複雌で父系の単位集団をもち、メスが集団間移籍をするという「構造」をもつが、この構造については社会生態学や霊長類社会学から説明がなされてきた。
社会生態学は、チンパンジー社会の「オスの集合性」と「メスの単独性」を対置する (Wrangham, 1987)。メスにとっての最重要資源である食物資源(果実)の分布様式への適応として、メスの単独性と狭いコアエリアの利用が説明され、そうしたメスが食物資源防衛と子殺しリスク低減のためにオスを集団内に囲い込む、とされる。一方オスは、オスにとっての最重要資源である繁殖資源(メス)の分布様式への適応として、オス同士が高い集合性を維持し、隣接他集団からメスを共同で防衛する、と説明される。この社会生態学によるチンパンジーの社会構造の説明では、オスの単独性という現象自体が問題にされていない。
霊長類社会学においても同様に、チンパンジー社会におけるオスの集合性に焦点をあてた議論が展開されている (黒田, 2009)。黒田 (2009)は、チンパンジーのオスは「優位志向者」であるとし、自らの順位を高めるためには他のオスとの連合が重要であり、またその連合関係の維持に常に気を配り確かめる必要からオス同士は高い集合性を維持する必要がある、と論じた。そのため、「チンパンジーのオスたちは孤立を恐れ、敵愾心を抑制し、関係確認の交渉を繰り返す」こと、また本発表との関連では、「チンパンジーのオスが順位争いで負けたあと単独行動するのは「敗者の自由」に近い」とまで述べている (黒田, 2009)。つまり、優位志向者であるチンパンジーのオスには「集団化の必然性」があり、「優位」が成立する条件としてチンパンジーのオスは集まらなければならず、優位であるメリットの対象であるメスを集めなければならない、というのである (黒田, 2009)。
しかし、これらはチンパンジーのオスの生活史をかなり単純化して捉えているのではないだろうか?例えば、タンザニア・ゴンベのチンパンジーの社交性の性差について、グドール (1986)は「オスの単独性」と「メスの非単独性(高い社交性)」について論じている。また同じくグドール (1986)はオスの優位志向性についても、すべてのオスが自らの社会的地位の上昇を目指すわけではなく、ごく短期間しか努力しないオスやすぐに順位上昇をあきらめてしまうオス、さらには自分の社会的地位にまったく関心を払っていないようにみえるオスについても言及し、「チンパンジーのオス=優位志向者」という単純な図式におさまらないオスの多様なあり方について論じている。
そこで、本発表では、タンザニア・マハレでこれまで観察されたオスのチンパンジーの単独生活の事例を検討することを通して、「移籍の可能性がなく」「高い集合性をもつ」とされるチンパンジーのオスの行動様式に「可能態(潜勢態)としての孤独」が含まれていることを示し、さらにその「可能態としての孤独」が社会にもたらすものについて議論する。
■タンザニア・マハレのチンパンジーのオスの単独行動事例
●集団からの長期的離脱→単独行動の例
・アルファオス交代にともなう集団からの長期的離脱の事例は、マハレではK集団で1例 (Nishida, 1983; 西田, 1981)とM集団で3例 (Uehara et al., 1994a; 上原, 1994; 保坂 and 西田, 2002; 西江, 2013, 2016)、タンザニアのゴンベ (Goodall, 1986, 1992)でも詳細に観察されているが、それほど頻繁に起こるわけではない。また、アルファオス以外の個体の長期的離脱・単独行動の事例もマハレでは2例観察されている (Nishida et al., 1995; Uehara et al., 1994b, 1994a, 上原, 1994, 2003)。また、マハレとゴンベ以外ではオスの長期的離脱や単独行動についての報告はない。
■考察
●チンパンジーのオスの単独生活を「可能態としての孤独」として見る
移籍の可能性が実質的にないと考えられるチンパンジーのオトナオスの単独生活は、ある意味では「出口なし」の極限的状態と考えられる。その最たる例が、K集団消滅後も長期にわたって単独での生活を余儀なくされたリモンゴの例といえるだろう。また、カジュギのように集団に復帰することなく消失した例もあり、チンパンジーのオスにとって単独生活から集団への復帰は容易ではないことが推察される。
ところがそうした「極限的状態」であると考えられるにも関わらず、実際に観察された単独生活中のオスのチンパンジーの様子は意外にも「普通」であり、おそらく際立った変化があまり見られないため過去の報告にも記載が少ない。単独生活中のカソンタを観察した西田 (1981)の記述や、単独生活中のカジュギの様子の記述 (上原, 1994; 西田, 1981)、また私自身が単独生活中のファナナを観察した印象としても (西江, 2016)、単独生活を送っているときのオスたちの姿は、離脱前と比べてもほとんど変化がなく、健康に過ごしている様子が報告されている。
このように、チンパンジーのオスは単独生活中でもそれ以前ととくに変わりなく「普通に」生活することが可能で、しかもそうした単独生活を数カ月から数年にわたって継続することすら可能である。チンパンジーのオスは「いつもそうする」わけではないが、「必要なときには集団から離脱して単独に近い生活を長期にわたって維持できる」という意味で、「潜勢態としての能力ないし性向」(内堀, 2009)としての孤独を、その行動様式のバリエーションのなかに備えているということができるだろう。こうした「可能態(潜勢態)としての孤独」をチンパンジーのオスの行動様式として描くことは、集団への「復帰」や優位志向性を前提としたり単独生活を「敗者の自由」(黒田, 2009)として描いたりするようなこれまでのチンパンジーのオスのステレオタイプとは異なる、新たな「チンパンジーのオスの可能態(潜勢態)としての社会性」を指し示しているといえるだろう。この「可能態(潜勢態)としての孤独」が社会にもたらしうるものの広がりを検討することで、社会構造の変化や単位集団の分裂・再編成の可能性、社会構造の進化史的祖型からの単独を含む小集団の析出の可能性、複数のコミュニティへの個体の帰属の可能性など、社会の柔軟性や新たな構造/非構造への移行といった「集団性の解体・再編成」の契機を探ることができると考えられる。
■文献
- Goodall, J. (1986). The Chimpanzees of Gombe: Patterns of Behavior (Cambridge, Mass: Belknap Press).
- Goodall, J. (1992). Unusual violence in the overthrow of an alpha male chimpanzee at Gombe. In Topics in Primatology vol.1: Human Origins, T. Nishida, W.C. McGrew, P. Marler, M. Pickford, and F.B.M. de Waal, eds. pp. 131–142.
- Itoh, N., and Nishida, T. (2007). Chimpanzee grouping patterns and food availability in Mahale Mountains National Park, Tanzania. Primates 48, 87–96.
- Nishida, T. (1968). The social group of wild chimpanzees in the Mahali Mountains. Primates 9, 167–224.
- Nishida, T. (1983). Alpha status and agonistic alliance in wild chimpanzees (Pan troglodytes schweinfurthii). Primates 24, 318–336.
- Nishida, T., Hosaka, K., Nakamura, M., and Hamai, M. (1995). A within-group gang attack on a young adult male chimpanzee: Ostracism of an ill-mannered member? Primates 36, 207–211.
- Uehara, S., Hiraiwa-Hasegawa, M., Hosaka, K., and Hamai, M. (1994a). The fate of defeated alpha male chimpanzees in relation to their social networks. Primates 35, 49–55.
- Uehara, S., Nishida, T., Takasaki, H., Norikoshi, K., Tsukahara, T., Nyundo, R., and Hamai, M. (1994b). A lone male chimpanzee in the wild: The survivor of a disintegrated unit-group. Primates 35, 275–281.
- Wrangham, R.W. (1987). Evolution of social structure. In Primate Societies, B.B. Smuts, R.M. Seyfarth, and R. Wrangham, eds. (Chicago: University Of Chicago Press), pp. 282–298.
- 上原重男 (1994). マハレ山塊国立公園で観察されたチンパンジーの雄の単独生活. 霊長類研究 10, 281–288.
- 上原重男 (2003). マハレのチンプ(ん?)紹介・第2回:リモンゴ. マハレ珍聞(マハレ野生動物保護協会ニューズレター) 2, 6.
- 伊谷純一郎 (1987). 霊長類社会の進化 (平凡社).
- 伊谷純一郎 (1993). 社会の構造と非構造. In 自然がほほ笑むとき, (平凡社), pp. 187–217.
- 保坂和彦, and 西田利貞 (2002). オストラシズム:アルファ雄、村八分からの復権. In マハレのチンパンジー:“パンスロポロジー”の三七年, 西田利貞, 川中健二, and 上原重男, eds. (京都大学学術出版会), pp. 439–471.
- 内堀基光 (2009). 単独者の集まり:孤独と「見えない」集団の間で. In 集団―人類社会の進化, 河合香吏, ed. (京都: 京都大学学術出版会), pp. 23–38.
- 河合香吏 (2009). 集団―人類社会の進化 (京都: 京都大学学術出版会).
- 西江仁徳 (2013). アルファオスとは「誰のこと」か?:チンパンジー社会における「順位」の制度的側面. In 制度―人類社会の進化, 河合香吏, ed. (京都: 京都大学学術出版会), pp. 121–142.
- 西江仁徳 (2016). 続・アルファオスとは「誰のこと」か?:チンパンジー社会における「他者」のあらわれ. In 他者―人類社会の進化, 河合香吏, ed. (京都: 京都大学学術出版会), pp. 125–148.
- 西田利貞 (1981). 野生チンパンジー観察記 (中央公論新社).
- 足立薫 (2009). 非構造の社会学:集団の極相へ. In 集団―人類社会の進化, 河合香吏, ed. (京都: 京都大学学術出版会), pp. 3–21.
- 黒田末寿 (2009). 集団的興奮と原始的戦争:平等原則とは何ものか?. In 集団―人類社会の進化, 河合香吏, ed. (京都: 京都大学学術出版会), pp. 255–274.
3.「今後の研究会活動に関するミーティング」(全員)
次期AA研・共同研究課題の申請および科学研究費補助金・財団助成金の申請について,代表の河合から話題提起をし,全員で議論した。
4.「チンパンジーは雑草種か?:西アフリカの農業景観における人との共存」(山越言)
ヒトの生息環境の極限条件を知るためには、ヒトに最も近縁な現生種チンパンジーの生息環境の多様性を理解し、そのなかでも極限的な生息状況に注目することは有益であろう。皮肉なことに、現代世界ではヒトとチンパンジーの生息環境は大きく重なり、チンパンジーにとって農業等の人為的影響を受けたいわゆる二次的植生が、極限的な生息地として重要になってきている。本発表では、野生チンパンジーを、原生息地である熱帯林に生息する「森の動物」と人為的環境に生息する「里の動物」とに区別する論考の系譜をまとめ、そのような視点が人と野生動物の共存にどのような示唆を与えるのかを考察する。
A. コルトラント(1986)は、西アフリカの野生チンパンジーのナッツ割り文化の起源として、森林由来のものと人里由来の二元論を主張し、人里由来の行動を些末で取るに足らないものと考える論者と激しい論争を行った。この古い論争が下敷きにしている、西アフリカ森林地域の人里環境、つまりは焼畑耕作と休閑地の植生がもたらすモザイク状で多様な農業景観は、有用樹アブラヤシを基盤とし、チンパンジーをはじめとする野生動物に豊かな食物資源を提供している。アブラヤシを半栽培的、またコモンズとして緩やかに管理し、野生動物による食害にも無頓着な現在の地域住民による資源管理法は興味深いが、アブラヤシの考古学的研究からは、火による景観管理に媒介された人びととアブラヤシとの「共生関係」が、3000-4000年の長い歴史を持つことも示唆されている。
少なくとも、西アフリカにおいて人為的景観管理が環境に爪痕を残すここ数千年のあいだ、人里近くの人為的環境は野生動物にとって新しいニッチとして存在し、そのようなニッチを好む「雑草種」を誘引してきたのではないだろうか。今日、人里近くに静かに「潜伏」し、アブラヤシに依存しながら焼畑による人為的景観を生息地として生き延びる西チンパンジーの生息状況は散発的に報告されているが、その実態は未知のままである。チンパンジーを神聖なものとして保護するギニア・ボッソウ村で観察される人為環境下のチンパンジーの生態や人為的環境への適応の姿は、そのような未知の「最後の類人猿」の生存を理解するための良い入り口になり得るだろう。
A. コルトラント(1986)は、西アフリカの野生チンパンジーのナッツ割り文化の起源として、森林由来のものと人里由来の二元論を主張し、人里由来の行動を些末で取るに足らないものと考える論者と激しい論争を行った。この古い論争が下敷きにしている、西アフリカ森林地域の人里環境、つまりは焼畑耕作と休閑地の植生がもたらすモザイク状で多様な農業景観は、有用樹アブラヤシを基盤とし、チンパンジーをはじめとする野生動物に豊かな食物資源を提供している。アブラヤシを半栽培的、またコモンズとして緩やかに管理し、野生動物による食害にも無頓着な現在の地域住民による資源管理法は興味深いが、アブラヤシの考古学的研究からは、火による景観管理に媒介された人びととアブラヤシとの「共生関係」が、3000-4000年の長い歴史を持つことも示唆されている。
少なくとも、西アフリカにおいて人為的景観管理が環境に爪痕を残すここ数千年のあいだ、人里近くの人為的環境は野生動物にとって新しいニッチとして存在し、そのようなニッチを好む「雑草種」を誘引してきたのではないだろうか。今日、人里近くに静かに「潜伏」し、アブラヤシに依存しながら焼畑による人為的景観を生息地として生き延びる西チンパンジーの生息状況は散発的に報告されているが、その実態は未知のままである。チンパンジーを神聖なものとして保護するギニア・ボッソウ村で観察される人為環境下のチンパンジーの生態や人為的環境への適応の姿は、そのような未知の「最後の類人猿」の生存を理解するための良い入り口になり得るだろう。
5.「人間仕掛けのロボット:日本古典芸能における二足直立と歩行の極限化」(船曳建夫)
人類が二足直立・歩行をいまだ完成させていないことは、頸椎から脊椎に至る箇所に疾患が絶えないことから推測される。また、本来、二足直立は、物理的に「安定」を取りにくい不利があることは、三足、四足のテーブルはあっても二足のテーブルがないことからそれは直感的に理解できよう。それにもかかわらず、人類は、脊椎動物で稀な、常時二足直立・歩行を行っていることは、「手」がもたらす有利性が、その不具合の不利を補ってあまりあるからと考えられる。本来、二足直立・歩行がもたらす有利性は、1)視界をより広く、遠くに取れる、2)立ち上がることによって身体を大きく見せ、相手を威嚇できる、3)前肢を手として道具使用に使える、の三点に絞ることが出来るが、特にその第三の点が、人類の文化・社会的進化を促すことがらとして、圧倒的な利点であると考えられるのだ。しかし、この発表では、その利点が人類の文化・社会的進化にどう関わってきたかについては論じない。むしろ、その有利さゆえに選択された二足直立・歩行が持つ「不利」、本来的な「不安定」が文化の身体表現にどのように関わっているか、ということについて考えたい。
この考察を始める契機となったのは、日本の身体を用いた芸能、パフォーマンスでは、二足直立・歩行の不安定の克服が、逆に「見せ場」として使われているということの気付きである。身体表現として、スポーツでは、オリンピックのモットーにもあるように、より高く、より早く、より強く、と身体の「動作」の極限を追い求めている。しかるに、日本列島に見られる芸能の伝統では、身体の「静止」の極限が求められている点が、対比的、逆説的に思え、興味を引く。
以下、その諸点を指摘する。
能楽においては、静止がそのパフォーマンスにおいて以下に大事であるか、は、演者が能舞台の上で、長時間の静止を行っているところから感得できる。シテの演技の始まりと終わり、その間においても、立った姿勢、座位の静止が数分から数十分の長さで行われる。ワキでは、その座位の静止の方が、舞台上の動作よりも長いことが多い。その静止が、演技の放棄であると誤解しやすいが、その静止にはある意図ともくろみがあるので、それは演技である。また、能楽における二足歩行では、上下動、横揺れが、抑制され、二足歩行がもたらす不安定が極力避けられている。また、静止においても移動においても、姿勢が、脊椎のS字形ではなく、「猫背」とも呼びうる、あえていえば、類人猿の立ち姿に近い、身体への負担が少なく、長時間同一の姿勢が取りやすいものになっていることは、興味深い。
人形浄瑠璃の人形の演技でも、主要な役柄は、静止の姿勢が重要な演技となる。不断に動く人形は、端役である。しかし、人形が動いていないとは、すなわち人形遣いが動いていないのである。本来胴体から足まで貫く軸がなく、二足では直立できない人形を、舞台上で、生きたものにするためには、人形遣いが静止をし、人形が二足直立をしているかのような静止をさせなければならない。二足の不安定を持つ人形遣いが、二足すら持たぬ人形に、二足の不安定さを与えた上で、それを二足の安定に持ち込む、という、アクロバティックな演技を行っているのである。
かぶきでは、二足直立・歩行の、集約された演技として「見得」がある。舞台のドラマが高調したときに、演技者は、その役柄の性根を表す姿形を取り、静止を維持する。その二足の姿勢に安定があることが求められる。しかし、それは、動作の放棄ではなく、動きから動きのあいだの静止である。そうした見得を言い表すには、その静止には動きがある、という矛盾したことばでするしかない。
こうした日本列島の芸能の身体技術は、一方で、他の文化の身体パフォーマンスにも見られることに留意したい。激しい超人的な身体運動、ムーブメント(movement)がその核心にあると見える西洋クラシックバレエでも、静止が重要な演技であるし、より高く飛び、より早く回り、より遠くに跳躍したときにも、最後は揺るがない静止として演技が締めくくられなければならない。また、現代にジャンルが確立された日本の舞踏(ぶとう)でも、能楽や歌舞伎に見られる静止や、移動が見られる。このジャンルの創始者である土方巽の言葉として伝えられる「舞踏とは命がけで突っ立った死体」とは、人類の文化表現としての二足直立・歩行を考えるに示唆的である。
本発表は、今後、日本の芸能における二足直立・歩行が、ロボットの二足直立・歩行を思わせるものであることを明らかにし、現代のロボットがある面で人間をモデルとしていることの逆に、日本列島では人間が「仮想上のロボット(人形)」を究極として表現を磨いてきた、興味深い事例の、人類の進化における意味を探り、今後の人間の身体性のあり方を予測する。
この考察を始める契機となったのは、日本の身体を用いた芸能、パフォーマンスでは、二足直立・歩行の不安定の克服が、逆に「見せ場」として使われているということの気付きである。身体表現として、スポーツでは、オリンピックのモットーにもあるように、より高く、より早く、より強く、と身体の「動作」の極限を追い求めている。しかるに、日本列島に見られる芸能の伝統では、身体の「静止」の極限が求められている点が、対比的、逆説的に思え、興味を引く。
以下、その諸点を指摘する。
能楽においては、静止がそのパフォーマンスにおいて以下に大事であるか、は、演者が能舞台の上で、長時間の静止を行っているところから感得できる。シテの演技の始まりと終わり、その間においても、立った姿勢、座位の静止が数分から数十分の長さで行われる。ワキでは、その座位の静止の方が、舞台上の動作よりも長いことが多い。その静止が、演技の放棄であると誤解しやすいが、その静止にはある意図ともくろみがあるので、それは演技である。また、能楽における二足歩行では、上下動、横揺れが、抑制され、二足歩行がもたらす不安定が極力避けられている。また、静止においても移動においても、姿勢が、脊椎のS字形ではなく、「猫背」とも呼びうる、あえていえば、類人猿の立ち姿に近い、身体への負担が少なく、長時間同一の姿勢が取りやすいものになっていることは、興味深い。
人形浄瑠璃の人形の演技でも、主要な役柄は、静止の姿勢が重要な演技となる。不断に動く人形は、端役である。しかし、人形が動いていないとは、すなわち人形遣いが動いていないのである。本来胴体から足まで貫く軸がなく、二足では直立できない人形を、舞台上で、生きたものにするためには、人形遣いが静止をし、人形が二足直立をしているかのような静止をさせなければならない。二足の不安定を持つ人形遣いが、二足すら持たぬ人形に、二足の不安定さを与えた上で、それを二足の安定に持ち込む、という、アクロバティックな演技を行っているのである。
かぶきでは、二足直立・歩行の、集約された演技として「見得」がある。舞台のドラマが高調したときに、演技者は、その役柄の性根を表す姿形を取り、静止を維持する。その二足の姿勢に安定があることが求められる。しかし、それは、動作の放棄ではなく、動きから動きのあいだの静止である。そうした見得を言い表すには、その静止には動きがある、という矛盾したことばでするしかない。
こうした日本列島の芸能の身体技術は、一方で、他の文化の身体パフォーマンスにも見られることに留意したい。激しい超人的な身体運動、ムーブメント(movement)がその核心にあると見える西洋クラシックバレエでも、静止が重要な演技であるし、より高く飛び、より早く回り、より遠くに跳躍したときにも、最後は揺るがない静止として演技が締めくくられなければならない。また、現代にジャンルが確立された日本の舞踏(ぶとう)でも、能楽や歌舞伎に見られる静止や、移動が見られる。このジャンルの創始者である土方巽の言葉として伝えられる「舞踏とは命がけで突っ立った死体」とは、人類の文化表現としての二足直立・歩行を考えるに示唆的である。
本発表は、今後、日本の芸能における二足直立・歩行が、ロボットの二足直立・歩行を思わせるものであることを明らかにし、現代のロボットがある面で人間をモデルとしていることの逆に、日本列島では人間が「仮想上のロボット(人形)」を究極として表現を磨いてきた、興味深い事例の、人類の進化における意味を探り、今後の人間の身体性のあり方を予測する。
第7回研究会での発表内容
1.「人新世という極限」(竹ノ下祐二)
生物の生存環境の極限としての人新世 (Anthropocene)
Crutzen & Stoermer (2000) は、完新世 (Holocene) は18世紀半ばに終了し、現代はあらたな地質時代に入ったとして、それを人新世 (Anthropocene)と呼ぶことを提唱した。彼らによれば、産業革命以降、人間活動の影響は地球の隅々まで及び、今や人間活動が地球規模の物質循環システムを変更するとともに、地球史上5回目の生物大量絶滅時代をもたらしているという。
あらゆる生物は環境と相互作用しながら生きており、その相互作用を通じて自らが生息環境から影響を受ける一方、自らも生息環境に影響を与え、多少なりともそれを改変する。生物によっては、その生存が生息環境に与える影響は極めて微々たるもので、ほとんど一方的に環境から影響を受けるばかりなこともある。それに対して、現代人による環境改変のインパクトは凄まじく、原生代における好気性細菌シアノバクテリアによる大気組成の大改編とそれに伴う生物の大量絶滅に匹敵するほどである。われわれを取り巻く環境の殆どはそれとわからなくても人為の影響を受けたものであり、このように自らによって改変された環境に生きるという状態は、それが人類自身の生存を脅かすということをさておいても、生物の生存環境の極限状態といえるだろう。
人新世をもたらしたヒトの特性
シアノバクテリアによる地球環境の大改編は、彼らが獲得した酸素呼吸という新しい能力によってもたらされた。では、ヒトによる現在の大規模環境改変を可能にしたのは、ホモ・サピエンスという種が獲得したどのような能力であろうか。 Crutzen & Stoermer (2000) は、先行研究を引用しながら、それはヒトが獲得した意識と思考力に基づく「思考の世界」であるという。ヒトは「今・ここ」の現実に基づき頭のなかで世界のモデルを構想し、それを操作することで未来予測をする。さらに、言語によってその思考世界を他者と共有することで、予測を現実化するために協力して環境改変を行うのである。
これは、私が人類社会の進化史的基盤研究(3)研究会において提唱した、「いま・ここ」の自分を離脱した超越的(演出家的)視点にもとづく「物語」を構築しそれを他者と共有して生きているという考え (竹ノ下 2016) に合致する。つまり、ヒトは物語を生きることによって未曾有の大繁栄を遂げ、ついには人新世という極限的時代を作り上げたといえる。
もう一つ、現在の人口増大を支えるヒトの生物学的特性として、繁殖に関してK戦略とrとを両立させていることがある。一般的に、生物の繁殖における子への投資においては「数」と「手厚さ」のトレードオフが見られる。すなわち、生物は「たくさん産む」か「手厚く育てるか」を選択しなくてはならない。だがヒトは協同育児によって「たくさん産んで、大事に育てる」戦略を取ることになった。
人新世のパラドクスとHomo sapiensの未来志向性
だが、超越的視点に基づくヒトの卓越した未来予測能力が人新世の到来を招いたというのは逆説的である。人類の生存に適した安定した環境を作るために行ってきた数々の環境改変は、結果として地球の歴史上まれに見るほどの不安定かつ非持続的な環境を作り上げてしまった。人為の影響が強い環境ほど、物理的、化学的、社会的、心理的にヒトの生存に適さないという状況が生じている。この逆説はどのように理解すればよいのだろうか。
そもそも、単に超越的視点を獲得したからといって、それを環境改変に用いなくてはならないという必然性はない。超越的視点によって「今・ここ」にとらわれない世界観を獲得したヒトが、「今・ここ」から離脱することを志向したことを説明する別の論理が必要である。
その論理は「物語の虚構性がもたらす不安と楽観」であると私は考える。まず、ヒトが「今・ここ」を離脱した超越的視点を獲得したといっても、それは所詮は「今・ここ」から再帰的、帰納的に得られた虚構にすぎない。その真実性は常に疑いにさらされており、生の実践を通じた現実とのマッチングと、他者とのコミュニケーションを通じた皆が同じ物語を共有していることの確認を通じて、不断に確認作業が続けられなくてはならない。物語の真実性への疑いは、その物語に基づいて作られた「今・ここ」の正当性への疑義となり、「このままでよいのか」と現状への不安を掻き立てるであろう。その一方で、ヒトは「今・ここ」から再帰的、帰納的に仮構した物語を、あたかも本当の超越的視点から眺めたものであるかのごとく理解する。それは、実際には不確かな未来予測であっても、それが超越的視点からもたらされた真実のシナリオだと信じさせる、未来に対する楽観的な態度を生み出すであろう。このようにして生み出された現在への不安と未来への楽観が、ヒトの未来に向けた環境改変への志向性をもたらしたのではないか。
未来志向性の進化史を考える
だが、ここでそうした未来志向性の起源や進化を考えようとすると、果たしてそれがヒト (Homo sapiens) 特有のものであり、人において初めて出現したといえるのかどうか、疑問が生じる。確かに、Homo sapiensは極めて未来志向性、フロンティア指向性の強い生物種で、それ以前の人類がなし得なかったような、「今・ここ」からは見えない陸地を目指して海に繰り出すことさえ行うようになった (海部, 2016)。だが、Homo sapiens以前の人類の進化史が分布拡大の歴史であったことを考えると、こうした「今・ここ」からの離脱への志向性の起源はもっと古いのではなかろうか。現代人の有りようのみに注目し、それを支える心的特性を考察するだけでは、本研究会の目標である「人類社会の進化史的基盤」を明らかにすることにはならない。ヒトが進化の過程でいつどのようにして現在の未来志向性につながる心的社会的特性を形成してきたのか、進化モデルではなく、ただ一度だけ起きた現実の人類進化史の年表を作る作業が必要である。それは、私が提唱した人類における他者性の進化モデルの各ステージを現実の人類進化史にあてはめてゆく作業でもある。
人類の進化史における形態、認知、社会進化の〈ずれ〉をどう説明するか
当然ながら、本発表でその年表を提示することは到底不可能である。そこで本発表では、今後の「年表づくり」に向けた課題としてふたつの観点を提示したい。その一つが、実際の人類進化における形態 (特に脳容量) の進化と認知および社会進化の“ずれ”である。
ふたたび人新世に話を戻せば、それは産業革命の始まった18世紀以降、ここ200年程度の出来事である。人類による大規模な環境改変のルーツを農耕の始まりに求めるとしても、それでもHomo sapiensの誕生からずっとあと、約1万年前のことである。
逆に時代を遡ると、人類初の石器文化であるオルドワン石器文化を形成したHomo habilisの脳容量は現生のチンパンジーや祖先種であるAustralopithecus afarensisと大差ない。しかし、一見シンプルに見えるオルドワン石器でも、その製作技術が広く伝播するには少なくとも gestural な教示行為が要求されるという研究もある (Morgan et al. 2010)。 “チンパンジー並の脳みそ” しかないHomo habilisがすでにそれだけの認知能力と協力性とを獲得していたとするならば、人新世の到来をHomo sapiensの高度な認知特性に帰することの根拠が揺らいでしまう。
さらに時代を遡ると、440万年前のArdipithecus ramidusでは、オスがメスに食物を運搬し分配していた可能性があるという (Lovejoy 2016)。しかし、やはり “チンパンジー並の脳みそ” しか持たない彼らが、チンパンジーの肉食の際の食物分配のような「今・ここ」における分配ならばともかく、「今・ここ」にいない個体に思いを馳せ、その個体に分配するために食物を運搬する、あるいは逆にその個体が食物を運んできてくれることを期待して待つ、などということを本当にしていたのか、疑問である。だが脳容量が小さい一方で、Ardipithecus ramidusは男女の体格の性差が小さく、またオスの犬歯も縮小しているなど、男女の間に穏やかで協力的な関係が成立していた可能性を示唆する形態学的特徴があることも事実である。
ヒトと大型類人猿の認知特性は質的に異なるのか?
もう一つの観点は、ヒトと大型類人猿の認知特性は本当に質的に異なるのか、という点である。ヒトは特別なのか、それともサルとの連続性のうちにあるのか、という議論は古くて新しい、そして不毛な議論である (プレマック 2017)。そして、大型類人猿の研究の歴史は、ヒトにしかないと考えられていた心理社会的能力や特性の多くが、程度や頻度の違いこそあれ、大型類人猿にも見られることを明らかにしてきた歴史でもある。例えば心の理論については、全くヒトと同様ではないにせよチンパンジーも他者の心を推測する事が明らかになった (Call & Tomasello 2008)。教示行為 teaching はヒト特有の現象と言われてきたが、近年チンパンジーの母子間で教示行為が見られたという報告があった (Musgrave et al. 2016)。西田はチンパンジーのアルファオスが「今・ここ」にいないライバルたちがよそでこっそり集まって毛づくろいしていると想像し、実際に集まって毛づくろいをしていたライバルたちを探し出して邪魔をした事例を報告している (西田 1994)。
ハビトゥスとしての超越的視点と未来志向性
もしかすると「「今・ここ」を離脱した超越的視点」は、ヒトでしか持ち得ないものではなく、せいぜいチンパンジー程度の脳があれば可能なことかもしれない。ただチンパンジーや化石人類、そして農耕開始以前のヒトはそれをごくまれに、いわば必殺技のように用いてきたのに対し、農耕開始以降、ヒトは次第にそれを日常的に用いるようになったということかもしれない。このように考えると、人新世をもたらしたヒトの超越的視点と未来志向性は、Homo sapiensという生物種に特有の能力の産物というより、むしろハビトゥス (ブルデュー 1995) と考えたほうがよいのかもしれない。だとするならば、その進化史をあとづけるには、メスーディらの文化進化論 (メスーディ 2016) や、ボウルズらによる協力性の進化に関する論考 (ボウルズ & ギンタス 2017) を参照しつつ論じてゆくのが有効であろう。
文献
Crutzen & Stoermer (2000) は、完新世 (Holocene) は18世紀半ばに終了し、現代はあらたな地質時代に入ったとして、それを人新世 (Anthropocene)と呼ぶことを提唱した。彼らによれば、産業革命以降、人間活動の影響は地球の隅々まで及び、今や人間活動が地球規模の物質循環システムを変更するとともに、地球史上5回目の生物大量絶滅時代をもたらしているという。
あらゆる生物は環境と相互作用しながら生きており、その相互作用を通じて自らが生息環境から影響を受ける一方、自らも生息環境に影響を与え、多少なりともそれを改変する。生物によっては、その生存が生息環境に与える影響は極めて微々たるもので、ほとんど一方的に環境から影響を受けるばかりなこともある。それに対して、現代人による環境改変のインパクトは凄まじく、原生代における好気性細菌シアノバクテリアによる大気組成の大改編とそれに伴う生物の大量絶滅に匹敵するほどである。われわれを取り巻く環境の殆どはそれとわからなくても人為の影響を受けたものであり、このように自らによって改変された環境に生きるという状態は、それが人類自身の生存を脅かすということをさておいても、生物の生存環境の極限状態といえるだろう。
人新世をもたらしたヒトの特性
シアノバクテリアによる地球環境の大改編は、彼らが獲得した酸素呼吸という新しい能力によってもたらされた。では、ヒトによる現在の大規模環境改変を可能にしたのは、ホモ・サピエンスという種が獲得したどのような能力であろうか。 Crutzen & Stoermer (2000) は、先行研究を引用しながら、それはヒトが獲得した意識と思考力に基づく「思考の世界」であるという。ヒトは「今・ここ」の現実に基づき頭のなかで世界のモデルを構想し、それを操作することで未来予測をする。さらに、言語によってその思考世界を他者と共有することで、予測を現実化するために協力して環境改変を行うのである。
これは、私が人類社会の進化史的基盤研究(3)研究会において提唱した、「いま・ここ」の自分を離脱した超越的(演出家的)視点にもとづく「物語」を構築しそれを他者と共有して生きているという考え (竹ノ下 2016) に合致する。つまり、ヒトは物語を生きることによって未曾有の大繁栄を遂げ、ついには人新世という極限的時代を作り上げたといえる。
もう一つ、現在の人口増大を支えるヒトの生物学的特性として、繁殖に関してK戦略とrとを両立させていることがある。一般的に、生物の繁殖における子への投資においては「数」と「手厚さ」のトレードオフが見られる。すなわち、生物は「たくさん産む」か「手厚く育てるか」を選択しなくてはならない。だがヒトは協同育児によって「たくさん産んで、大事に育てる」戦略を取ることになった。
人新世のパラドクスとHomo sapiensの未来志向性
だが、超越的視点に基づくヒトの卓越した未来予測能力が人新世の到来を招いたというのは逆説的である。人類の生存に適した安定した環境を作るために行ってきた数々の環境改変は、結果として地球の歴史上まれに見るほどの不安定かつ非持続的な環境を作り上げてしまった。人為の影響が強い環境ほど、物理的、化学的、社会的、心理的にヒトの生存に適さないという状況が生じている。この逆説はどのように理解すればよいのだろうか。
そもそも、単に超越的視点を獲得したからといって、それを環境改変に用いなくてはならないという必然性はない。超越的視点によって「今・ここ」にとらわれない世界観を獲得したヒトが、「今・ここ」から離脱することを志向したことを説明する別の論理が必要である。
その論理は「物語の虚構性がもたらす不安と楽観」であると私は考える。まず、ヒトが「今・ここ」を離脱した超越的視点を獲得したといっても、それは所詮は「今・ここ」から再帰的、帰納的に得られた虚構にすぎない。その真実性は常に疑いにさらされており、生の実践を通じた現実とのマッチングと、他者とのコミュニケーションを通じた皆が同じ物語を共有していることの確認を通じて、不断に確認作業が続けられなくてはならない。物語の真実性への疑いは、その物語に基づいて作られた「今・ここ」の正当性への疑義となり、「このままでよいのか」と現状への不安を掻き立てるであろう。その一方で、ヒトは「今・ここ」から再帰的、帰納的に仮構した物語を、あたかも本当の超越的視点から眺めたものであるかのごとく理解する。それは、実際には不確かな未来予測であっても、それが超越的視点からもたらされた真実のシナリオだと信じさせる、未来に対する楽観的な態度を生み出すであろう。このようにして生み出された現在への不安と未来への楽観が、ヒトの未来に向けた環境改変への志向性をもたらしたのではないか。
未来志向性の進化史を考える
だが、ここでそうした未来志向性の起源や進化を考えようとすると、果たしてそれがヒト (Homo sapiens) 特有のものであり、人において初めて出現したといえるのかどうか、疑問が生じる。確かに、Homo sapiensは極めて未来志向性、フロンティア指向性の強い生物種で、それ以前の人類がなし得なかったような、「今・ここ」からは見えない陸地を目指して海に繰り出すことさえ行うようになった (海部, 2016)。だが、Homo sapiens以前の人類の進化史が分布拡大の歴史であったことを考えると、こうした「今・ここ」からの離脱への志向性の起源はもっと古いのではなかろうか。現代人の有りようのみに注目し、それを支える心的特性を考察するだけでは、本研究会の目標である「人類社会の進化史的基盤」を明らかにすることにはならない。ヒトが進化の過程でいつどのようにして現在の未来志向性につながる心的社会的特性を形成してきたのか、進化モデルではなく、ただ一度だけ起きた現実の人類進化史の年表を作る作業が必要である。それは、私が提唱した人類における他者性の進化モデルの各ステージを現実の人類進化史にあてはめてゆく作業でもある。
人類の進化史における形態、認知、社会進化の〈ずれ〉をどう説明するか
当然ながら、本発表でその年表を提示することは到底不可能である。そこで本発表では、今後の「年表づくり」に向けた課題としてふたつの観点を提示したい。その一つが、実際の人類進化における形態 (特に脳容量) の進化と認知および社会進化の“ずれ”である。
ふたたび人新世に話を戻せば、それは産業革命の始まった18世紀以降、ここ200年程度の出来事である。人類による大規模な環境改変のルーツを農耕の始まりに求めるとしても、それでもHomo sapiensの誕生からずっとあと、約1万年前のことである。
逆に時代を遡ると、人類初の石器文化であるオルドワン石器文化を形成したHomo habilisの脳容量は現生のチンパンジーや祖先種であるAustralopithecus afarensisと大差ない。しかし、一見シンプルに見えるオルドワン石器でも、その製作技術が広く伝播するには少なくとも gestural な教示行為が要求されるという研究もある (Morgan et al. 2010)。 “チンパンジー並の脳みそ” しかないHomo habilisがすでにそれだけの認知能力と協力性とを獲得していたとするならば、人新世の到来をHomo sapiensの高度な認知特性に帰することの根拠が揺らいでしまう。
さらに時代を遡ると、440万年前のArdipithecus ramidusでは、オスがメスに食物を運搬し分配していた可能性があるという (Lovejoy 2016)。しかし、やはり “チンパンジー並の脳みそ” しか持たない彼らが、チンパンジーの肉食の際の食物分配のような「今・ここ」における分配ならばともかく、「今・ここ」にいない個体に思いを馳せ、その個体に分配するために食物を運搬する、あるいは逆にその個体が食物を運んできてくれることを期待して待つ、などということを本当にしていたのか、疑問である。だが脳容量が小さい一方で、Ardipithecus ramidusは男女の体格の性差が小さく、またオスの犬歯も縮小しているなど、男女の間に穏やかで協力的な関係が成立していた可能性を示唆する形態学的特徴があることも事実である。
ヒトと大型類人猿の認知特性は質的に異なるのか?
もう一つの観点は、ヒトと大型類人猿の認知特性は本当に質的に異なるのか、という点である。ヒトは特別なのか、それともサルとの連続性のうちにあるのか、という議論は古くて新しい、そして不毛な議論である (プレマック 2017)。そして、大型類人猿の研究の歴史は、ヒトにしかないと考えられていた心理社会的能力や特性の多くが、程度や頻度の違いこそあれ、大型類人猿にも見られることを明らかにしてきた歴史でもある。例えば心の理論については、全くヒトと同様ではないにせよチンパンジーも他者の心を推測する事が明らかになった (Call & Tomasello 2008)。教示行為 teaching はヒト特有の現象と言われてきたが、近年チンパンジーの母子間で教示行為が見られたという報告があった (Musgrave et al. 2016)。西田はチンパンジーのアルファオスが「今・ここ」にいないライバルたちがよそでこっそり集まって毛づくろいしていると想像し、実際に集まって毛づくろいをしていたライバルたちを探し出して邪魔をした事例を報告している (西田 1994)。
ハビトゥスとしての超越的視点と未来志向性
もしかすると「「今・ここ」を離脱した超越的視点」は、ヒトでしか持ち得ないものではなく、せいぜいチンパンジー程度の脳があれば可能なことかもしれない。ただチンパンジーや化石人類、そして農耕開始以前のヒトはそれをごくまれに、いわば必殺技のように用いてきたのに対し、農耕開始以降、ヒトは次第にそれを日常的に用いるようになったということかもしれない。このように考えると、人新世をもたらしたヒトの超越的視点と未来志向性は、Homo sapiensという生物種に特有の能力の産物というより、むしろハビトゥス (ブルデュー 1995) と考えたほうがよいのかもしれない。だとするならば、その進化史をあとづけるには、メスーディらの文化進化論 (メスーディ 2016) や、ボウルズらによる協力性の進化に関する論考 (ボウルズ & ギンタス 2017) を参照しつつ論じてゆくのが有効であろう。
文献
- ブルデュー P, 1993.『資本主義のハビトゥス―アルジェリアの矛盾』 藤原書店.
- Call J, Tomasello M, 2008. Does the chimpanzee have a theory of mind? 30 years later. Trends in Cognitive Sciences, 12:187-192.
- Crutzen PJ, Stoermer EF, 2000. The “Anthropocene”. Global Change Newsletter, 41:17–18.
- ボウルズ S, ギンタス H, 2017.『協力する種―制度と心の共進化』 NTT出版.
- 海部陽介, 2016.『日本人はどこから来たのか?』 文藝春秋.
- Lovejoy O, 2009. Reexamining human origins in light of Ardipitecus ramidus. Science, 326(2): 74–74e8.
- メスーディ A, 2016.『文化進化論―ダーウィン進化論は文化を説明できるか』 NTT出版.
- Morgan TJH, Uomini NT, Rendell RE, Chouinard-Thuly L, Street SE, Lewis HM, Cross CP, Evans C, Kearney R, de la Torre I, Whiten A, Laland KN, 2010. Experimental evidence for the co-evolution of hominin tool-making teaching and language. Nature Communications 6.
- Musgrave S, Morgan D, Lonsdorf E, Mundry R, Sanz C, 2016. Tool transfers are a form of teaching among chimpanzees. Scientific Reports, 6:34783.
- プレマック D, 2017.『ギャバガイ! ―「動物のことば」の先にあるもの』勁草書房.
- 竹ノ下祐二, 2016. 「社会という『物語』―分業,協同育児と他者性の進化」,河合香吏(編), 『他者―人類社会の進化』京都大学学術出版会. pp.379–388.
2.「生きる世界の極―死者と精霊そして放射能の手がかりに」(西井凉子)
触知できない存在、「今・ここ」にない存在にむきあう営為は、いかにも人間特有の創造力や感受性によっているかのうように思われる。しかし、はたしてそうであろうか。兆候や不安、恐怖、目に見えないものを感じる能力、それに何らかの反応をすることは人間以外にはないのであろうか。
私たちの生きている現実は、身体の感覚器官による認識に限られない。そもそも、見るといった行為さえも「見える」から「見る」にかわることで、ある対象が認識される。一方、五感といった感覚器官で捉えられない徴候は、通常は異界に存在するといわれる精霊だったり死者といった存在として行為遂行的に出現する。こうした五感を超えた徴候をよみとり、恐怖や期待を抱き、そしてそれによる行為をすることは、じつは人間以外の動物でもやっているのではないか。
では、人間特有の経験とは何か。それは、感受することもなく、身体的反応もないところで生命の危機にさらされるような経験を考えることができる。福島の原発事故により放射線にさらされることは、まさにこうした経験であるといえる。放射線は「測る」ことによってしか検知できず、その身体への影響は甚大である。「測る」ことによって出現した現実は、五感によって世界を感知し生きる現実と齟齬をきたす。それは、通常は感じられず、考慮にも入れられない生命の土台となるような空気だったり大地だったりするもので、人間と人間以外のすべての生命に共通する生の潜在性ともいえる。そこに変化があるときにはじめて、われわれの生活のなかに出現し、見えるようになり、対処すべきものになるのではないか。人間以外の生命は、ただ身体として影響をうけ死ぬ、もしくは生き延びる。しかし、人間のみが、それを科学技術によって触知可能として、身体を超えたところで現実を作りだす。
まとめよう。本発表では、目に見えない、触知できない精霊や死者といったきわめて人間特有の文化的作為にみえるものも、身体で感受することから兆候をよみとり対応しようとする人間以外の霊長類を含む動物にも共通する可能性があるとした。身体を離脱して現実を創造する技術のみが人間特有の経験を作り出すといえるかもしれない。
私たちの生きている現実は、身体の感覚器官による認識に限られない。そもそも、見るといった行為さえも「見える」から「見る」にかわることで、ある対象が認識される。一方、五感といった感覚器官で捉えられない徴候は、通常は異界に存在するといわれる精霊だったり死者といった存在として行為遂行的に出現する。こうした五感を超えた徴候をよみとり、恐怖や期待を抱き、そしてそれによる行為をすることは、じつは人間以外の動物でもやっているのではないか。
では、人間特有の経験とは何か。それは、感受することもなく、身体的反応もないところで生命の危機にさらされるような経験を考えることができる。福島の原発事故により放射線にさらされることは、まさにこうした経験であるといえる。放射線は「測る」ことによってしか検知できず、その身体への影響は甚大である。「測る」ことによって出現した現実は、五感によって世界を感知し生きる現実と齟齬をきたす。それは、通常は感じられず、考慮にも入れられない生命の土台となるような空気だったり大地だったりするもので、人間と人間以外のすべての生命に共通する生の潜在性ともいえる。そこに変化があるときにはじめて、われわれの生活のなかに出現し、見えるようになり、対処すべきものになるのではないか。人間以外の生命は、ただ身体として影響をうけ死ぬ、もしくは生き延びる。しかし、人間のみが、それを科学技術によって触知可能として、身体を超えたところで現実を作りだす。
まとめよう。本発表では、目に見えない、触知できない精霊や死者といったきわめて人間特有の文化的作為にみえるものも、身体で感受することから兆候をよみとり対応しようとする人間以外の霊長類を含む動物にも共通する可能性があるとした。身体を離脱して現実を創造する技術のみが人間特有の経験を作り出すといえるかもしれない。
3.「ハザード状況下における環境と生存に関する試論―フィリピン南部の紛争と福島原発事故・放射能災害の事例から」(床呂郁哉)
本報告では極限的なハザード(hazard)状況下における生存と環境の関係に関する試論的考察を行った。事例としてはフィリピン南部における「モロ」(Moro:フィリピンのムスリム少数民族)をめぐる紛争・暴力、そして東日本大震災に伴う福島原発事故による放射能災害の二つの事例を取り上げて、そうした状況に巻き込まれた避難民の生存と環境の問題を試論的に考察していく。その際の参照枠組みとしては複数の環境のレイヤーをめぐる議論(河合)が参考となりうると考える。すなわち自然(生態)環境、社会環境、象徴(情報)環境というアイディアを参照しながら、その相互作用のイフェクトとしてハザードを捉えることを検討した。
まず本報告で言うハザードとは何を意味するのか、その定義について述べた。一般的にはハザードは地震や津波・台風などによる自然災害などを指すことが多い(「例:ハザードマップ」)が、本報告では必ずしもそれだけには限定されず、むしろ人間社会の側の要因が絡む出来事、とくに紛争やテロ事件等なども含まれる概念として規定した。
報告前半ではフィリピン南部におけるモロ(Moro)人の社会をめぐる紛争・暴力の事例を検討した。最初にフィリピン南部における近年の紛争・暴力をめぐる状況の概要等について述べた後で特に今年(2017年)5月から10月までミンダナオ島のマラウィ市で発生したテロ組織IS(Islamic Stateイスラム国)系の武装集団(Maute Groupeマウテ・グループ)による同市の占領事件と、その結果としての大量の国内避難民(IDP)発生の状況について述べた。そして報告者によるIDPへの現地での聞き取り調査に基づき極限としての紛争状況について検討した。
報告の後半では東日本大震災に伴う福島第一原子力発電所の事故とその結果としての放射能災害の事例を取り上げた。そこでは同原発事故による避難民らの語りの紹介と分析を通じて、原発事故・災害の特徴に関して検討した。概して福島原発事故では環境内に放出された大量の放射性物質とその長期に渡る影響が問題となる。その影響は人間による直接の暴力・テロであるとか自然災害でも津波被害などと異なり、その被害や影響が短期的には見えにくいというある種の不可視性を帯びているが、その不可視の放射能と、その情報環境内での可視化がこの種の災害では重要であることを指摘した。すなわち、同事故の影響では、自然環境内での放射性物質の「客観的」挙動だけではなく、それをどう計測し伝達し、いかに評価し意味づけするのか、というリスクに関する情報の解釈や受容のされ方の差異によってその後の被災者の行動や生活パターンに影響してくることを情報環境という概念などを用いながら分析を試みた。
まず本報告で言うハザードとは何を意味するのか、その定義について述べた。一般的にはハザードは地震や津波・台風などによる自然災害などを指すことが多い(「例:ハザードマップ」)が、本報告では必ずしもそれだけには限定されず、むしろ人間社会の側の要因が絡む出来事、とくに紛争やテロ事件等なども含まれる概念として規定した。
報告前半ではフィリピン南部におけるモロ(Moro)人の社会をめぐる紛争・暴力の事例を検討した。最初にフィリピン南部における近年の紛争・暴力をめぐる状況の概要等について述べた後で特に今年(2017年)5月から10月までミンダナオ島のマラウィ市で発生したテロ組織IS(Islamic Stateイスラム国)系の武装集団(Maute Groupeマウテ・グループ)による同市の占領事件と、その結果としての大量の国内避難民(IDP)発生の状況について述べた。そして報告者によるIDPへの現地での聞き取り調査に基づき極限としての紛争状況について検討した。
報告の後半では東日本大震災に伴う福島第一原子力発電所の事故とその結果としての放射能災害の事例を取り上げた。そこでは同原発事故による避難民らの語りの紹介と分析を通じて、原発事故・災害の特徴に関して検討した。概して福島原発事故では環境内に放出された大量の放射性物質とその長期に渡る影響が問題となる。その影響は人間による直接の暴力・テロであるとか自然災害でも津波被害などと異なり、その被害や影響が短期的には見えにくいというある種の不可視性を帯びているが、その不可視の放射能と、その情報環境内での可視化がこの種の災害では重要であることを指摘した。すなわち、同事故の影響では、自然環境内での放射性物質の「客観的」挙動だけではなく、それをどう計測し伝達し、いかに評価し意味づけするのか、というリスクに関する情報の解釈や受容のされ方の差異によってその後の被災者の行動や生活パターンに影響してくることを情報環境という概念などを用いながら分析を試みた。
4.「チンパンジー社会における共在の極限:新入りメスのふるまいに着目して」(花村俊吉)
1.はじめに:チンパンジー社会における共在の諸相と新入りメス
野生チンパンジーは複雄複雌の集団を形成し、その構成個体が出会いと別れを繰り返す(離合集散)。2個体が別れてから次に再会するまでの時間間隔は数分のこともあれば数ヵ月に及ぶこともあり、個体間の距離も数キロになることがある。こうした離合集散の過程で、チンパンジーは視覚的に接触しうるそのつどの個体たちの集まりである「対面パーティ」を形成する。さらに、タンザニアのマハレM集団では、頻繁な離合集散や声を介して1日の遊動方向を概ね同調させる個体たちのゆるやかなまとまりである「遊動パーティ」が観察され、そのパーティサイズは主食である果実の密度に応じて大きくかつコンスタントに季節変動する[Itoh and Nishida 2007;Hanamura 2015a]。このようにチンパンジーは、集団の他個体との共在/非共在状態を、視覚・聴覚の双方を介して、時間的・空間的にさまざまなスケールにおいて日々経験している。
これまでの研究から、M集団のチンパンジーたちが、長距離音声・パントフートを鳴き交わすことで非対面下の出会い(互いに「相手が自分に気づいている」と期待しうる状態)を達成していること、しかしその後どのように相互行為を続けるかという「ゴール」は決まっておらず、それぞれの生活リズムや視界内他個体との関わりの影響も受けつつ、互いに相手の出方次第という「プロセス」志向的な態度で行為を調整していることがわかっている。こうしたやり方によって、彼らは、声さえ届くならいつでも出会い試みたり共在状態の継続を試みたりすることを可能にすると同時に、いつでもそうした相互行為を中断してそれぞれの活動を継続したり、互いの声を聴き流してそのまま離れるに任せたりすることもまた可能にしていると考えられる[花村 2013]。また、M集団のチンパンジーたちは、稀に聴こえてくるよそ者(他集団個体)の声に対しても基本的にはプロセス志向的な態度で接するが、そのリアクションは知り合い(自集団他個体)の声に対するそれとは大きく異なる。その違いを分析することで、彼らにとってのよそ者と知り合いの位置づけを考察することができる。よそ者は、非敵対的に共在したことがないがゆえに、その声が聴こえてきたときに、「何が起こるかわからない」出会いの可能性に備えて「離れていることのできない(次の声を待ったり他の痕跡を探したりし続けることになる)」他者である。それに対して、知り合いは、非敵対的な共在を繰り返してきた結果、その声が聴こえてきたとしても、「これまでと同じであろう」出会いの可能性にとくに備えることなく「離れていることのできる」他者である。しかしこの差異は、チンパンジーが言語をもたず、「経験=実際の相互行為」に先立ってこと・ものを概念化し共有する術をもたないであろうことを踏まえると、それぞれとの相互行為の積み重ねの有無・多寡によって生じているはずであり、その意味で結果として大きな断絶があるのだが、あくまで連続的なものだと考えられる[花村 2016]。
このような社会において、数十から百頭ほどのチンパンジーたちが、人間の言語のような手がかりも、ニホンザルの空間的な近接(群れ)のような手がかりもなしに、結果として非敵対的な共在を繰り返してきたということのみを根拠に「同じ集団のメンバーであること(あり続けること)」を実現している[伊藤 2006, 2016;北村 2016を参照]。そこにはどのような共在/非共在の工夫―「別れと再会」の反復を可能にする機序―がみられるのか。この課題にアプローチするために、本発表では、集団に移入して間もない個体のふるまいに着目した。
チンパンジー社会では、未経産の若いメスが集団間を移出入し、そうしたメスは、原則、出産を迎えた集団に生涯留まる[Nishida and Kawanaka 1972;Nakamura 2015]。そこで本発表では、移入したあと出産を迎えるまでの「新入りメス」が、移入先集団における馴染みのない社会的・生態的環境を身体化しつつ生存していく―「集団のメンバーになる」―過程で、「既住個体」とどのような関係を築き、どのような共在/非共在状態を経験しているのかを検討した。まず、新入りメスと既住個体の関係についての通説と近年のマハレM集団におけるその実態を紹介した。そのうえで、新入りメスが既住個体と「はぐれる」場面(視覚的・聴覚的な共在の切れ目・極限)―常に母親と遊動する未成熟な個体を除けば、チンパンジーにとって他個体と別れることは常態であり、それによって慌てることは基本的にはない―や、パントフートを介した相互行為における(既住個体と比べた・在住年数に応じた)新入りメスのふるまいの特徴を分析し、「集団のメンバーであること」が他個体とどのように共在/非共在することであるかを逆照射することを目指した。なお、新入りメスは、とくに移入直後、観察者である人間を恐れるので観察が難しい部分があるが、人間の存在に馴れるということもヒトづけの進んだM集団のメンバーになることの一部である。
2.新入りメスと既住個体の関係
自然選択(性選択)の帰結として、哺乳類のメスは、食物資源を確保することに、オスは繁殖機会を増加することに重きを置いた行動傾向をもつように進化してきたとされる。この仮定に基づけば、既住メスは、食い扶持を減らしうる新入りメスに対して威嚇・攻撃しやすく、オスは、そうした新入りメスと他個体の敵対的交渉において、繁殖機会を増やしうる新入りメスを応援・保護しやすく、それゆえ新入りメスも既住メスを避けオスと遊動することが多いと予測される。これらの予測を支持する結果が、マハレM集団や他地域の複数の集団で報告されている[Nishida 1989;Pusey 1980;Kahlenberg et al. 2008a, b;Pusey and Schroepfer-Walker 2013]。
しかし、近年のマハレM集団の新入りメスと既住個体の相互行為についての調査から、これらの通説とは異なる以下の結果が得られている[Hanamura 2015bも参照]
1)新入りメスは、既住メスと比べて単独(ただし既住メスの大部分は子どもを連れている)で過ごす時間が短い傾向があり、オスだけでなく、オスと同じくらい既住メスとも一緒にいる。
2)少なくとも一部の新入りメスは、特定の高齢の既住メス(以下「共遊動メス」)にくっていて遊動することがあり、その関係は出産後も長期的に継続する可能性がある。
3)新入りメスは、既住メスよりもむしろオスから威嚇・攻撃されることが多く、既住メスの威嚇・攻撃の多くは、採食場面ではなく、アカンボウやコドモ、あるいはその母親である既住メスに対する新入りメスの関心の高さがきっかけとなって生じる。
4)新入りメスと他個体との敵対的交渉時に、オスだけでなく既住メスも新入りメスを応援・保護することがあり、それらの既住メスは「共遊動メス」に限定されない。
したがって、通説とは異なり、新入りメスの移入過程において、オスとの関係だけでなく、メス、とくに既住メスとの関係も等しく重要であると考えられる。そのため、以下の新入りメスの既住個体との共在/非共在のあり方に関する分析では、相手の性別によらず分析を進める。
なお、通説とは異なる結果が得られた背景には、調査時の新入りメスの数や特徴(年齢や体重、個性、それまでの移出入経験の有無など)、集団のサイズや社会性比、あるいは食物の種類や量・分布の違いなど、複数の要因が存在すると考えられる。たとえば、マハレの先行研究[Nishida 1989]は、隣接するK集団が消失する過程でメスが多数移入してきていた1980年代前半のM集団でおこなわれた研究であり、その新入りメスの多くが通常の未経産メスとは異なる、一度K集団に定着したあと二次的にM集団に移入してきた経産メスであったということが、結果に影響しているだろう。
3.新入りメスの他個体とはぐれたときのふるまいとパントフートを介した相互行為の特徴
M集団の新入りメスを調査してきて観察した事例と、パントフートを介した相互行為に関して得た量的なデータから、以下のことが示唆される。
1)移入したての、とくに年齢が若めの新入りメスは、オスメス含めたさまざまな既住個体に(しかも同時に)毛づくろいされまくることがあるなど人気があり、新入りメスもそのつどさまざまな既住個体への追随を試みるが、その過程でどこにどんな採食樹があるか、また場合によっては何が食べられるものであるかを知ることになる(こういった社会的環境と生態的環境の重なりは、私がチンパンジーを追跡することで日々生じることでもある)。ただし、新入りメスの追随に対してその相手は、追随を許容すれども新入りメスを慮って待ったりはしないし、新入りメスの入念な毛づくろいの後に毛づくろいし返さなかったりもする。
2)新入りメスは、それまでに一緒にいた他個体が視界内からいなくなって単独になったことに気づいたとき、既住個体では滅多にみられないような慌てぶりを示すことがある(こうした場面を「別れる」ではなく「はぐれる」と表現することができる)。新入りメスにとって、遠くから聴こえてくる声より目の前にいる他個体の方が大事であり、新入りメスはそのつど一緒にいる視界内他個体の動向をかなり気にしているが、たとえば採食に夢中になっていてはぐれることがある。
3)しかし新入りメスは、はぐれたとしても困ってどうしようもなくなるというわけではなく、しばらくすると落ち着き、はぐれたならそれはそれで採食・休息などしつつ、のんびりと誰かが通りかかるのを待ったり、声が聴こえてくるのを待ったりする。他個体とはぐれたときに、パントフートを始発して返事があるか試したり、聴こえてきたパントフートに返事して相手に自分の位置や存在を知らせたりするなど、新入りメスもパントフートを介した相互行為を試みたりおこなったりすることがあるが、とくに在住年数の短い新入りメスは、在住年数の長い新入りメスと比べてパントフートの始発を開始する頻度が低い傾向がある。パントフートの始発は常に返事を期待して発されるわけはないが[花村 2013]、この傾向の背景には、移入して間もない新入りメスにとっては、その場所で始発して「誰」に聴かれうるかよくわからない、たとえ返事があってもまだ「誰」からの返事かよくわからないといった理由があるかもしれない。また、サンプル数が少ないため正確な評価は困難だが、移入して間もない新入りメスはパントフートの被返事率も低い可能性がある。これが事実であれば、そこには、既住個体に声を覚えられていない、返事されるほどには既住個体との関係が深まっていない、返事がありそうな文脈がよくわからないといった理由が考えられるし、被返事率が低いということ自体も始発頻度が低くなる理由になりうる。そういった状況に置かれていることもあって、新入りメスは(とくに在住年数がある程度長くなってくると)パントフートを「試し打ち」して返事があるかどうか試すこともあるようだ。
4)新入りメスの観察時には既住メスの観察時より視界外からパントフートが聴こえてくる頻度が高い傾向がある。この結果は、新入りメスが他個体とはぐれたときに、声の聴こえそうな範囲に留まるように自身の遊動ルートを調整している可能性を示唆する。新入りメスは既住メスに比べて、パントフートが聴こえてきたときにその声の方に走って向かうことが多いという結果も、この解釈と矛盾しない。
このように、新入りメスは、既住メスと比べて他個体との視覚的・聴覚的な共在状態を維持・継続しようとする傾向が強い。その背景には、しばらくすると落ち着きを取り戻していたようにはぐれても何とかなるのだが(何せ彼女らは他集団からたいてい1頭でやって来るのだし、再移出することもあるのだ)、はぐれたら食物の探索に手間がかかるという生態的な理由のほか、今後その個体たちと―ひいてはその集団の他の個体たちとも―再会できるかどうかわからないという社会的な理由が存在すると考えられる。
4.考察(今後の課題)
4−1.新入りメスの他個体との共在の諸相
チンパンジーの聴覚的な共在に関するこれまでの私の研究では、パントフートを介して非対面下の出会いに至るやり方や過程、非対面下で出会ったあとの相互行為のやり方や共在の度合いの変容過程に着目してきた。しかし、出会いには至らずとも声を聴いて遊動方向を調整したり、声の方に走っていったりという微妙な状態もまたある。それは、マハレのチンパンジー社会、とくに連日のように大きな「遊動パーティ」が形成される集合季にはかなり常態的な状態である。そして、その一方で集合季であってもそうした状態から抜けて単独ないし少数個体のみとで遊動することが常に可能である既住個体と比べて、新入りメスは、そうした状態を維持・継続する(声の聴こえる範囲に留まるよう遊動方向を調整する)ことへのモチベーションが高いようだ。つまり、新入りメスにとって、移入先集団の他個体は、視覚的・聴覚的な共在をまだあまり繰り返してきていないため、既住個体にとっての集団内他個体よりも「離れていることのできない」他者である。その声を聴いて関心や好奇心が喚起されていると考えられるので、緊張や不安が喚起される「よそ者」とはベクトルは異なるが、あまり「離れていることができない」という点で、「よそ者」と「知り合い」の中間的な存在として立ち現れていると言えるだろう。
また、新入りメスはそのつどさまざまな個体を追随したり、個体によっては特定の既住メスを繰り返し追随したりしており、他個体との聴覚的な共在(新入りメスにはそれが難しいという側面もある)よりも視覚的な共在の維持・継続が重要であることが確認できた。とりわけ、声もあまり聴こえず、複数の小さな「遊動パーティ」がそれぞれ独立に遊動する分散季にはそうであると考えられるが、前者は後者の共在に比べて、相手が声をあげないとどうにもならないという意味で不確定要素が強いということがその理由のひとつだろう。そのため、視界内に誰もいなくなったこと、あるいは追随を繰り返してきた特定の共遊動メスがいなくなったことに気づいたときに慌てる(はぐれる)ことがあるのだと考えられる。今後、聴覚的な共在/非共在と視覚的なそれとの重なりと違いについて、さらに検討する必要がある。
4−2.「別れと再会」を可能にする機序
チンパンジーは、視覚的・聴覚的な共在を繰り返すことによって「同じ集団のメンバーであること」を不断に生成し続けていると考えられるが、今後この点について、新入りメスの他個体からはぐれたときのふるまいやパントフートを介した相互行為の特徴を、既住個体の他個体と別れるときのふるまいやパントフートを介した相互行為の特徴と比較しつつさらに理解を深めていく必要がある。大澤[2016]が「チンパンジーは他個体と別れても気にしない」と述べているように、人間の感覚からすると、チンパンジーの別れはびっくりするくらい淡泊にみえることも多いし、実際にしばしば「そのあと何ごともなかったかのように、その後の行動を続ける」[ibid. p176]。しかし、チンパンジーが離れゆく他個体のことを気にしていないわけでは決してないし、そうして「離れていることができる」ことにこそ一定数のチンパンジーたちが離合集散しつつ「同じ集団のメンバーであること」を実現している共在/非共在の工夫を理解する鍵があるはずである。また、別れても結果的に再会してきたということによって「同じ集団のメンバーであること」を根拠づけていると考えられるチンパンジーと、別れの挨拶(しばしば再会の約束を意味する言語が伴う)をおこなったり、「同じ集団のメンバーであること」を担保するさまざまな「物語」(制度)に依拠して再会を信じたり規範化したりする人間とを比較することで、人類がその進化史において、より広い範囲の個体間で「別れと再会」を繰り返し、時間的・空間的にもより長く・遠く「離れていることができる」ようになった機序を探ることもできるかもしれない。
引用文献
野生チンパンジーは複雄複雌の集団を形成し、その構成個体が出会いと別れを繰り返す(離合集散)。2個体が別れてから次に再会するまでの時間間隔は数分のこともあれば数ヵ月に及ぶこともあり、個体間の距離も数キロになることがある。こうした離合集散の過程で、チンパンジーは視覚的に接触しうるそのつどの個体たちの集まりである「対面パーティ」を形成する。さらに、タンザニアのマハレM集団では、頻繁な離合集散や声を介して1日の遊動方向を概ね同調させる個体たちのゆるやかなまとまりである「遊動パーティ」が観察され、そのパーティサイズは主食である果実の密度に応じて大きくかつコンスタントに季節変動する[Itoh and Nishida 2007;Hanamura 2015a]。このようにチンパンジーは、集団の他個体との共在/非共在状態を、視覚・聴覚の双方を介して、時間的・空間的にさまざまなスケールにおいて日々経験している。
これまでの研究から、M集団のチンパンジーたちが、長距離音声・パントフートを鳴き交わすことで非対面下の出会い(互いに「相手が自分に気づいている」と期待しうる状態)を達成していること、しかしその後どのように相互行為を続けるかという「ゴール」は決まっておらず、それぞれの生活リズムや視界内他個体との関わりの影響も受けつつ、互いに相手の出方次第という「プロセス」志向的な態度で行為を調整していることがわかっている。こうしたやり方によって、彼らは、声さえ届くならいつでも出会い試みたり共在状態の継続を試みたりすることを可能にすると同時に、いつでもそうした相互行為を中断してそれぞれの活動を継続したり、互いの声を聴き流してそのまま離れるに任せたりすることもまた可能にしていると考えられる[花村 2013]。また、M集団のチンパンジーたちは、稀に聴こえてくるよそ者(他集団個体)の声に対しても基本的にはプロセス志向的な態度で接するが、そのリアクションは知り合い(自集団他個体)の声に対するそれとは大きく異なる。その違いを分析することで、彼らにとってのよそ者と知り合いの位置づけを考察することができる。よそ者は、非敵対的に共在したことがないがゆえに、その声が聴こえてきたときに、「何が起こるかわからない」出会いの可能性に備えて「離れていることのできない(次の声を待ったり他の痕跡を探したりし続けることになる)」他者である。それに対して、知り合いは、非敵対的な共在を繰り返してきた結果、その声が聴こえてきたとしても、「これまでと同じであろう」出会いの可能性にとくに備えることなく「離れていることのできる」他者である。しかしこの差異は、チンパンジーが言語をもたず、「経験=実際の相互行為」に先立ってこと・ものを概念化し共有する術をもたないであろうことを踏まえると、それぞれとの相互行為の積み重ねの有無・多寡によって生じているはずであり、その意味で結果として大きな断絶があるのだが、あくまで連続的なものだと考えられる[花村 2016]。
このような社会において、数十から百頭ほどのチンパンジーたちが、人間の言語のような手がかりも、ニホンザルの空間的な近接(群れ)のような手がかりもなしに、結果として非敵対的な共在を繰り返してきたということのみを根拠に「同じ集団のメンバーであること(あり続けること)」を実現している[伊藤 2006, 2016;北村 2016を参照]。そこにはどのような共在/非共在の工夫―「別れと再会」の反復を可能にする機序―がみられるのか。この課題にアプローチするために、本発表では、集団に移入して間もない個体のふるまいに着目した。
チンパンジー社会では、未経産の若いメスが集団間を移出入し、そうしたメスは、原則、出産を迎えた集団に生涯留まる[Nishida and Kawanaka 1972;Nakamura 2015]。そこで本発表では、移入したあと出産を迎えるまでの「新入りメス」が、移入先集団における馴染みのない社会的・生態的環境を身体化しつつ生存していく―「集団のメンバーになる」―過程で、「既住個体」とどのような関係を築き、どのような共在/非共在状態を経験しているのかを検討した。まず、新入りメスと既住個体の関係についての通説と近年のマハレM集団におけるその実態を紹介した。そのうえで、新入りメスが既住個体と「はぐれる」場面(視覚的・聴覚的な共在の切れ目・極限)―常に母親と遊動する未成熟な個体を除けば、チンパンジーにとって他個体と別れることは常態であり、それによって慌てることは基本的にはない―や、パントフートを介した相互行為における(既住個体と比べた・在住年数に応じた)新入りメスのふるまいの特徴を分析し、「集団のメンバーであること」が他個体とどのように共在/非共在することであるかを逆照射することを目指した。なお、新入りメスは、とくに移入直後、観察者である人間を恐れるので観察が難しい部分があるが、人間の存在に馴れるということもヒトづけの進んだM集団のメンバーになることの一部である。
2.新入りメスと既住個体の関係
自然選択(性選択)の帰結として、哺乳類のメスは、食物資源を確保することに、オスは繁殖機会を増加することに重きを置いた行動傾向をもつように進化してきたとされる。この仮定に基づけば、既住メスは、食い扶持を減らしうる新入りメスに対して威嚇・攻撃しやすく、オスは、そうした新入りメスと他個体の敵対的交渉において、繁殖機会を増やしうる新入りメスを応援・保護しやすく、それゆえ新入りメスも既住メスを避けオスと遊動することが多いと予測される。これらの予測を支持する結果が、マハレM集団や他地域の複数の集団で報告されている[Nishida 1989;Pusey 1980;Kahlenberg et al. 2008a, b;Pusey and Schroepfer-Walker 2013]。
しかし、近年のマハレM集団の新入りメスと既住個体の相互行為についての調査から、これらの通説とは異なる以下の結果が得られている[Hanamura 2015bも参照]
1)新入りメスは、既住メスと比べて単独(ただし既住メスの大部分は子どもを連れている)で過ごす時間が短い傾向があり、オスだけでなく、オスと同じくらい既住メスとも一緒にいる。
2)少なくとも一部の新入りメスは、特定の高齢の既住メス(以下「共遊動メス」)にくっていて遊動することがあり、その関係は出産後も長期的に継続する可能性がある。
3)新入りメスは、既住メスよりもむしろオスから威嚇・攻撃されることが多く、既住メスの威嚇・攻撃の多くは、採食場面ではなく、アカンボウやコドモ、あるいはその母親である既住メスに対する新入りメスの関心の高さがきっかけとなって生じる。
4)新入りメスと他個体との敵対的交渉時に、オスだけでなく既住メスも新入りメスを応援・保護することがあり、それらの既住メスは「共遊動メス」に限定されない。
したがって、通説とは異なり、新入りメスの移入過程において、オスとの関係だけでなく、メス、とくに既住メスとの関係も等しく重要であると考えられる。そのため、以下の新入りメスの既住個体との共在/非共在のあり方に関する分析では、相手の性別によらず分析を進める。
なお、通説とは異なる結果が得られた背景には、調査時の新入りメスの数や特徴(年齢や体重、個性、それまでの移出入経験の有無など)、集団のサイズや社会性比、あるいは食物の種類や量・分布の違いなど、複数の要因が存在すると考えられる。たとえば、マハレの先行研究[Nishida 1989]は、隣接するK集団が消失する過程でメスが多数移入してきていた1980年代前半のM集団でおこなわれた研究であり、その新入りメスの多くが通常の未経産メスとは異なる、一度K集団に定着したあと二次的にM集団に移入してきた経産メスであったということが、結果に影響しているだろう。
3.新入りメスの他個体とはぐれたときのふるまいとパントフートを介した相互行為の特徴
M集団の新入りメスを調査してきて観察した事例と、パントフートを介した相互行為に関して得た量的なデータから、以下のことが示唆される。
1)移入したての、とくに年齢が若めの新入りメスは、オスメス含めたさまざまな既住個体に(しかも同時に)毛づくろいされまくることがあるなど人気があり、新入りメスもそのつどさまざまな既住個体への追随を試みるが、その過程でどこにどんな採食樹があるか、また場合によっては何が食べられるものであるかを知ることになる(こういった社会的環境と生態的環境の重なりは、私がチンパンジーを追跡することで日々生じることでもある)。ただし、新入りメスの追随に対してその相手は、追随を許容すれども新入りメスを慮って待ったりはしないし、新入りメスの入念な毛づくろいの後に毛づくろいし返さなかったりもする。
2)新入りメスは、それまでに一緒にいた他個体が視界内からいなくなって単独になったことに気づいたとき、既住個体では滅多にみられないような慌てぶりを示すことがある(こうした場面を「別れる」ではなく「はぐれる」と表現することができる)。新入りメスにとって、遠くから聴こえてくる声より目の前にいる他個体の方が大事であり、新入りメスはそのつど一緒にいる視界内他個体の動向をかなり気にしているが、たとえば採食に夢中になっていてはぐれることがある。
3)しかし新入りメスは、はぐれたとしても困ってどうしようもなくなるというわけではなく、しばらくすると落ち着き、はぐれたならそれはそれで採食・休息などしつつ、のんびりと誰かが通りかかるのを待ったり、声が聴こえてくるのを待ったりする。他個体とはぐれたときに、パントフートを始発して返事があるか試したり、聴こえてきたパントフートに返事して相手に自分の位置や存在を知らせたりするなど、新入りメスもパントフートを介した相互行為を試みたりおこなったりすることがあるが、とくに在住年数の短い新入りメスは、在住年数の長い新入りメスと比べてパントフートの始発を開始する頻度が低い傾向がある。パントフートの始発は常に返事を期待して発されるわけはないが[花村 2013]、この傾向の背景には、移入して間もない新入りメスにとっては、その場所で始発して「誰」に聴かれうるかよくわからない、たとえ返事があってもまだ「誰」からの返事かよくわからないといった理由があるかもしれない。また、サンプル数が少ないため正確な評価は困難だが、移入して間もない新入りメスはパントフートの被返事率も低い可能性がある。これが事実であれば、そこには、既住個体に声を覚えられていない、返事されるほどには既住個体との関係が深まっていない、返事がありそうな文脈がよくわからないといった理由が考えられるし、被返事率が低いということ自体も始発頻度が低くなる理由になりうる。そういった状況に置かれていることもあって、新入りメスは(とくに在住年数がある程度長くなってくると)パントフートを「試し打ち」して返事があるかどうか試すこともあるようだ。
4)新入りメスの観察時には既住メスの観察時より視界外からパントフートが聴こえてくる頻度が高い傾向がある。この結果は、新入りメスが他個体とはぐれたときに、声の聴こえそうな範囲に留まるように自身の遊動ルートを調整している可能性を示唆する。新入りメスは既住メスに比べて、パントフートが聴こえてきたときにその声の方に走って向かうことが多いという結果も、この解釈と矛盾しない。
このように、新入りメスは、既住メスと比べて他個体との視覚的・聴覚的な共在状態を維持・継続しようとする傾向が強い。その背景には、しばらくすると落ち着きを取り戻していたようにはぐれても何とかなるのだが(何せ彼女らは他集団からたいてい1頭でやって来るのだし、再移出することもあるのだ)、はぐれたら食物の探索に手間がかかるという生態的な理由のほか、今後その個体たちと―ひいてはその集団の他の個体たちとも―再会できるかどうかわからないという社会的な理由が存在すると考えられる。
4.考察(今後の課題)
4−1.新入りメスの他個体との共在の諸相
チンパンジーの聴覚的な共在に関するこれまでの私の研究では、パントフートを介して非対面下の出会いに至るやり方や過程、非対面下で出会ったあとの相互行為のやり方や共在の度合いの変容過程に着目してきた。しかし、出会いには至らずとも声を聴いて遊動方向を調整したり、声の方に走っていったりという微妙な状態もまたある。それは、マハレのチンパンジー社会、とくに連日のように大きな「遊動パーティ」が形成される集合季にはかなり常態的な状態である。そして、その一方で集合季であってもそうした状態から抜けて単独ないし少数個体のみとで遊動することが常に可能である既住個体と比べて、新入りメスは、そうした状態を維持・継続する(声の聴こえる範囲に留まるよう遊動方向を調整する)ことへのモチベーションが高いようだ。つまり、新入りメスにとって、移入先集団の他個体は、視覚的・聴覚的な共在をまだあまり繰り返してきていないため、既住個体にとっての集団内他個体よりも「離れていることのできない」他者である。その声を聴いて関心や好奇心が喚起されていると考えられるので、緊張や不安が喚起される「よそ者」とはベクトルは異なるが、あまり「離れていることができない」という点で、「よそ者」と「知り合い」の中間的な存在として立ち現れていると言えるだろう。
また、新入りメスはそのつどさまざまな個体を追随したり、個体によっては特定の既住メスを繰り返し追随したりしており、他個体との聴覚的な共在(新入りメスにはそれが難しいという側面もある)よりも視覚的な共在の維持・継続が重要であることが確認できた。とりわけ、声もあまり聴こえず、複数の小さな「遊動パーティ」がそれぞれ独立に遊動する分散季にはそうであると考えられるが、前者は後者の共在に比べて、相手が声をあげないとどうにもならないという意味で不確定要素が強いということがその理由のひとつだろう。そのため、視界内に誰もいなくなったこと、あるいは追随を繰り返してきた特定の共遊動メスがいなくなったことに気づいたときに慌てる(はぐれる)ことがあるのだと考えられる。今後、聴覚的な共在/非共在と視覚的なそれとの重なりと違いについて、さらに検討する必要がある。
4−2.「別れと再会」を可能にする機序
チンパンジーは、視覚的・聴覚的な共在を繰り返すことによって「同じ集団のメンバーであること」を不断に生成し続けていると考えられるが、今後この点について、新入りメスの他個体からはぐれたときのふるまいやパントフートを介した相互行為の特徴を、既住個体の他個体と別れるときのふるまいやパントフートを介した相互行為の特徴と比較しつつさらに理解を深めていく必要がある。大澤[2016]が「チンパンジーは他個体と別れても気にしない」と述べているように、人間の感覚からすると、チンパンジーの別れはびっくりするくらい淡泊にみえることも多いし、実際にしばしば「そのあと何ごともなかったかのように、その後の行動を続ける」[ibid. p176]。しかし、チンパンジーが離れゆく他個体のことを気にしていないわけでは決してないし、そうして「離れていることができる」ことにこそ一定数のチンパンジーたちが離合集散しつつ「同じ集団のメンバーであること」を実現している共在/非共在の工夫を理解する鍵があるはずである。また、別れても結果的に再会してきたということによって「同じ集団のメンバーであること」を根拠づけていると考えられるチンパンジーと、別れの挨拶(しばしば再会の約束を意味する言語が伴う)をおこなったり、「同じ集団のメンバーであること」を担保するさまざまな「物語」(制度)に依拠して再会を信じたり規範化したりする人間とを比較することで、人類がその進化史において、より広い範囲の個体間で「別れと再会」を繰り返し、時間的・空間的にもより長く・遠く「離れていることができる」ようになった機序を探ることもできるかもしれない。
引用文献
- 花村俊吉 2013 「見えない他者の声に耳を澄ませるとき:チンパンジーのプロセス志向的な慣習と制度の可能態」 河合香吏(編)『制度:人類社会の進化』 京都大学学術出版会, pp. 167–194.
- Hanamura S 2015a Fission-fusion grouping. In: M Nakamura, K Hosaka, N Itoh and K Zamma (eds) Mahale Chimpanzees: 50 Years of Research. Cambridge: Cambridge University Press, pp. 106–118.
- Hanamura S 2015b Male-female relationships: affiliative, interventional, and dominant-subordinate interactions. In: M Nakamura, K Hosaka, N Itoh and K Zamma (eds) Mahale Chimpanzees: 50 Years of Research. Cambridge: Cambridge University Press, pp. 410–420.
- 花村俊吉 2016 「見えないよそ者の声に耳を欹てるとき:チンパンジー社会における他者」 河合香吏(編) 『他者:人類社会の進化』 京都大学学術出版会, pp. 177–205.
- 伊藤詞子 2006 「まとまることのメカニズム」 西田正規・北村光二・山極寿一(編)『人間性の起源と進化』 昭和堂, pp. 233–262.
- Itoh N and Nishida T 2007 Chimpanzees grouping patterns and food availability in Mahale Mountains National Park, Tanzania. Primates 48: 87–96.
- Kahlenberg SM, Emery Thompson M, Muller MN and Wrangham RW 2008. Immigration costs for female chimpanzees and male protection as an immigrant counterstrategy to intrasexual aggression. Animal Behaviour 76: 1497–1509.
- Kahlenberg SM, Emery Thompson M and Wrangham RW 2008b Female competition over core areas in Pan troglodytes schweinfurthii, Kibale National Park, Uganda. International Journal of Primatology 29: 931–947.
- Nakamura M 2015 Demography of the M group. In: M Nakamura, K Hosaka, N Itoh and K Zamma (eds) Mahale Chimpanzees: 50 Years of Research. Cambridge: Cambridge University Press, pp. 82–93.
- Nishida T 1989 Social interactions between resident and immigrant female chimpanzees. In: PG Heltne, LA Marquardt (eds.) Understanding Chimpanzees, Cambridge, MA: Harvard University Press, pp. 68–89.
- Nishida T and Kawanaka K 1972 Inter-unit-group relationship among wild chimpanzees of the Mahali Mountains. Kyoto University African Studies 7: 131–169.
- 大澤真幸 2016 「〈社会性〉への不可能な進化」 『現代思想 vol. 44-10』 青土社, pp. 165–177.
- Pusey A 1980 Inbreeding avoidance in chimpanzees. Animal Behaviour 28: 543–552.
- Pusey A and Schroepfer-Walker K 2013 Female competition in chimpanzees. Philosophical Transactions of the Royal Society B 368: 20130077.
第8回研究会での発表内容
内容
成果論集の刊行に向けたミーティングをおこなった。具体的には成果論集(『極限—人類社会の進化』)に執筆予定者(本共同研究課題メンバーの全員。ただし、当日欠席の床呂郁哉と竹ノ下祐二は文書にて参加)が順次,自身の執筆予定の内容・要旨について報告し,全員で議論した。また,論集全体の目的・概要についても全員で議論した。